En el núcleo del espermatozoide, la cromatina se encuentra de 6 a 8 veces más condensada que en los núcleos de las células somáticas. Esta condensación es el resultado del intercambio de histonas por protaminas durante el proceso de formación de los espermatozoides, otorgando al material genético una resistencia única al ataque por especies reactivas de oxígeno, así como al estrés que implica el trayecto hasta su encuentro con el ovocito en el oviducto.
Esta condensación tan alta debe ser relajada, una vez producida la fecundación del ovocito, para permitir la formación del pronúcleo masculino y la posterior singamia. Para que ocurra la descondensación, es necesaria la reducción de los puentes disulfuro (S-S) de las protaminas, reacción que lleva a cabo el glutatión y, además, la remoción de las protaminas con un intercambio posterior o concomitante por las histonas ovocitarias. En este punto, todavía no hay consenso sobre la molécula involucrada.
Mecanismo Molecular de la Descondensación
Papel del Heparán Sulfato (HS)
Nuestro laboratorio estudia la descondensación nuclear del espermatozoide humano in vitro, intentando ahondar en la comprensión de las etapas del proceso y las moléculas involucradas en el mismo. La descondensación requiere la reducción de puentes disulfuro en las protaminas y la remoción de las mismas para ser reemplazadas por histonas ovocitarias. Los espermatozoides humanos pueden descondensarse in vitro con heparina y glutatión.
Estudios previos han observado que el heparán sulfato (HS), análogo estructural de la heparina, pero no así otros glicosaminoglicanos, presenta actividad descondensante in vitro similar a la heparina. Se ha propuesto que el HS actuaría in vivo como aceptor de protaminas.
Los objetivos de una investigación reciente fueron (1) acercarse a las condiciones in vivo analizando el comportamiento de núcleos aislados frente a la descondensación con heparina y glutatión in vitro y (2) demostrar que el HS está presente en el ovocito y podría funcionar como agente descondensante. Se utilizaron espermatozoides de donantes normospérmicos (OMS) y ovocitos de ratón.
Los núcleos espermáticos aislados descondensaron con glutatión y heparán sulfato o heparina, pero no con otros glicosaminoglicanos. El análisis por microscopía confocal reveló localización de HS en el citoplasma y el espacio perivitelino del ovocito. La utilización de enzimas específicas indicó que la capacidad descondensante del ovocito podía atribuirse a la presencia de HS. Estas son las primeras evidencias de la presencia de HS en el ovocito y de su posible rol como aceptor de protaminas durante la descondensación del espermatozoide humano in vivo.
Eventos Clave en la Descondensación
A diferencia de las células somáticas, el espermatozoide posee su cromatina empaquetada principalmente por protaminas en lugar de histonas. Las cisteínas presentes en las protaminas forman puentes disulfuro intra e intercatenarios, lo que le proporciona al núcleo espermático gran estabilidad y resistencia para poder atravesar el tracto masculino, el femenino y llegar con su ADN intacto hasta el sitio de fecundación.
Luego de que el espermatozoide penetra el ovocito, es imprescindible que se produzca la descondensación de su cromatina para que se reemplacen las protaminas espermáticas por histonas ovocitarias. La descondensación de la cromatina involucra dos eventos principales: se deben reducir los puentes disulfuro (tiorreducción) y removerse las protaminas. Las moléculas involucradas en estos procesos in vivo serían el glutatión reducido (GSH) y el heparán sulfato (HS) ovocitarios.
Importancia Clínica de la Descondensación de la Cromatina Espermática
Impacto en la Fertilidad y Reproducción Asistida
La integridad del genoma paterno es de vital importancia en el inicio y mantenimiento de un embarazo a término, tanto in vivo como in vitro. Espermatozoides con ADN dañado pueden fecundar ovocitos en metafase II con la misma eficacia que espermatozoides con ADN intacto.
Se estima que un 25% de las causas de infertilidad masculina son de origen idiopático. La etiología de algunas de estas causas podría estar relacionada con daño de ADN en los espermatozoides. Sin embargo, este daño podría ser reparado por el ovocito después de la fecundación. Esto va a depender, sobre todo, (i) de la calidad citoplasmática y genómica del ovocito, y (ii) del grado de daño en las cadenas de ADN del espermatozoide que haya fecundado al ovocito.
Dado que la capacidad del ovocito de reparar este tipo de daño disminuye con la edad y que, al mismo tiempo, el nivel de fragmentación de ADN en los espermatozoides aumenta, ello podría explicar, al menos en parte, la disminución significativa en la tasa de embarazo que se observa en parejas de edad avanzada. Este tipo de daño en las cadenas de ADN podría encontrarse en embriones con un complemento cromosómico normal.
Es decir, la presencia de daño de ADN no reparado por encima de un umbral crítico en embriones generados tanto in vivo como in vitro, podría explicar el paro en el desarrollo embrionario que se produce tras la implantación de embriones con un cariotipo normal.
Las técnicas de reproducción asistida han podido resolver problemas de motilidad, defectos espermáticos, concentración de gametos muy reducida en el fluido seminal (oligozoospermia), pero todavía no pueden solucionar los defectos de la descondensación del núcleo espermático dentro del ovocito. Esto pone en evidencia la importancia del proceso de descondensación, que no puede ser reemplazado por las técnicas de reproducción asistida.
✨🧬 Así es el proceso de FECUNDACIÓN - Fertilización Paso a paso
La Fragmentación del ADN Espermático
Cuando el proceso de condensación de la cromatina, que ocurre durante la espermatogénesis para mantener la integridad del espermatozoide, se ve afectado por diversas causas, puede resultar en fallas de la descondensación que se evidencian una vez que el ovocito es penetrado.
El ADN del espermatozoide aporta la mitad del material genético a la descendencia, y se requiere que sea sano para la fertilización, el desarrollo del embrión, el correcto desarrollo fetal y posnatal; de lo contrario, un ADN anormal altera cualquiera de estos procesos. Actualmente la infertilidad es un problema global que va en aumento y se ha demostrado que la calidad del ADN del espermatozoide puede afectar la fecundación.
Es por ello que la evaluación de la integridad del ADN, además del estudio de los parámetros seminales, puede aportar información adicional acerca de la calidad del espermatozoide, y ser de gran ayuda para identificar las causas de la infertilidad masculina y así orientar de mejor manera a las parejas en los centros de reproducción asistida, como el CREAFAM en Puebla, México.
Causas de la Fragmentación del ADN Espermático
El daño de ADN en los espermatozoides puede afectar tanto el ADN mitocondrial como el nuclear y puede ser inducido por cinco mecanismos principales:
- Apoptosis durante el proceso de espermatogénesis: Durante la espermatogénesis, un screening celular importante resulta en la inducción de apoptosis en un 50-60% de las células germinales que entran en la meiosis I. Estas células, marcadas con marcadores apoptóticos tipo Fas, deberían ser fagocitadas y eliminadas por la célula de Sertoli a la cual se encuentran asociadas. Sin embargo, esto no siempre ocurre, y un porcentaje variable de estas células germinales entra en el proceso de remodelado celular de la espermiogénesis, apareciendo posteriormente en el eyaculado. Un estudio reciente sugiere que existe una disociación entre la calidad genómica de la célula germinal y el remodelado espermático que tiene lugar durante la espermiogénesis. Esto significa que una célula germinal puede tener el núcleo “pulverizado” por apoptosis o ser aneuploide, y sin embargo el espermatozoide resultante puede tener una morfología normal. De ahí que cuando se microinyecte un espermatozoide de morfología normal, esto no garantiza que el genoma sea normal. Se ha constatado que cuando existe oligospermia, la probabilidad de que un espermatozoide con morfología normal sea aneuploide es mucho mayor que si existe normozoospermia, probablemente relacionado con un bloqueo madurativo parcial asociado a alteraciones meióticas. El hecho de que un porcentaje variable de espermatozoides en el eyaculado expresan marcadores apoptóticos como Fas, fosfatidilserina, Bcl-XL, p53, podría utilizarse para seleccionar espermatozoides no apoptóticos.
- Roturas de ADN o nicks producidos durante el remodelado de la cromatina: Alteraciones en el proceso de remodelado de la cromatina espermática durante la espermiogénesis podrían resultar en fragmentación de ADN. Se ha postulado que la presencia de roturas en el ADN podría ser indicativa de maduración incompleta durante la espermiogénesis. Para el empaquetamiento de la cromatina del espermatozoide, es necesaria la actividad de nucleasas endógenas que corten y liguen el ADN durante su protaminación. Alteraciones en el control de este proceso podrían resultar en roturas de ADN no reparadas.
- Fragmentación de ADN a nivel post-testicular: Estudios recientes demuestran que espermatozoides inmaduros que producen niveles elevados de especies reactivas de oxígeno (ROS) pueden inducir daño de ADN en espermatozoides maduros. Este daño se produciría después de la espermiación durante la co-migración de espermatozoides maduros e inmaduros desde los túbulos seminíferos al epidídimo. La corta vida media de los ROS facilitaría que estos induzcan fragmentación de ADN a nivel del epidídimo, ya sea actuando directamente sobre el ADN o indirectamente mediante la activación de endonucleasas y caspasas espermáticas. El epidídimo podría también jugar un papel activo en la inducción de fragmentación de ADN en los espermatozoides a su paso, ya sea por radicales libres (anión superóxido, radical hidroxilo o óxido nítrico) o mediante la activación de caspasas y endonucleasas espermáticas por agentes tóxicos o factores epididimarios. Este tipo de daño podría prevenirse con antioxidantes en el primer caso, pero no en el segundo.
- Fragmentación de ADN inducida por caspasas y endonucleasas espermáticas: La activación de caspasas y endonucleasas en espermatozoides diferenciados por factores físico-químicos puede inducir también fragmentación del ADN espermático. La exposición de espermatozoides de ratón in vitro a 40ºC o la exposición de los testículos a 42ºC in vivo, resulta en un aumento significativo en el grado de fragmentación del ADN.
- Fragmentación de ADN inducida por radio y quimioterapia.
Además, se sabe que la exposición a tóxicos ambientales puede provocar efectos deletéreos sobre la calidad de los espermatozoides y la fertilidad masculina. Esto permite pensar en utilizar la descondensación de la cromatina como un posible indicador del efecto de disruptores endocrinos, tanto in vivo como in vitro.
Evaluación de la Integridad del ADN Espermático
Técnicas Diagnósticas
Recientemente se han introducido una serie de tests para la determinación de daño de ADN en los espermatozoides. Un aspecto importante es el tipo de roturas que se produce en las cadenas de ADN (cadena sencilla versus cadena doble) y la susceptibilidad a la fragmentación de ADN.
Tests como el SCSA, SCD, o COMET a pH ácido o alcalino requieren un paso inicial de desnaturalización para detectar los fragmentos de ADN o roturas potenciales en la cadena de ADN. Cuando se observa daño de ADN en condiciones de pH ácido o alcalino y no en condiciones de pH neutro, se habla de puntos lábiles de rotura al tratamiento ácido o alcalino.
Por el contrario, el test TUNEL, ISNT y el COMET a pH neutro no requieren un paso previo de desnaturalización y miden roturas auténticas de ADN de cadena sencilla (ISNT, TUNEL y COMET) y cadena doble (TUNEL y COMET). Dado que el pH intracelular en el ovocito es de 7.0, roturas de ADN de cadena sencilla o la presencia de puntos lábiles de rotura en el ADN no tendrían mayores consecuencias en la formación del pronúcleo masculino, ya que a pH neutro las cadenas de ADN no se disocian y, por lo tanto, este tipo de daño sería mucho más fácil de reparar.
Por lo tanto, podrían considerarse dos tipos de tests:
- Tests que miden “daño real” de ADN, como TUNEL, ISNT o COMET a pH neutro.
- Tests que miden “daño potencial” de ADN o puntos lábiles de rotura al tratamiento ácido o alcalino y susceptibilidad a la desnaturalización de ADN, como SCSA, SCD, Chromomycin A3 y COMET a pH ácido o alcalino.
Los tests que miden daño real de ADN deberían tener un mayor valor predictivo.
Ensayo de Túnel (TUNEL)
Permite visualizar la incorporación de nucleótidos marcados en los extremos del ADN que quedan libres a causa de las rupturas, bien sean de cadena simple o doble. La señal es mayor cuanto mayor sea el grado de fragmentación del ADN. Esta técnica fue desarrollada para identificar una población apoptótica de espermatozoides en el eyaculado, sin embargo, la fragmentación del ADN medida mediante el TUNEL no está siempre asociada con un fenómeno apoptótico, ya que podría deberse a una maduración espermática defectuosa por un empaquetamiento incorrecto de la cromatina.
Ensayo In Situ Nick Translation (ISNT)
Cuantifica la incorporación dUTP biotinilado en las rupturas del ADN de cadena sencilla en una reacción catalizada por la polimerasa I. Detecta a los espermatozoides que contienen valores apreciables de daño endógeno en su ADN, indicando la presencia de anomalías originadas durante la remodelación de la cromatina del espermatozoide. El ISNT evidencia una buena correlación con la movilidad y la morfología espermática, y en menor medida con la concentración de espermatozoides en el semen.
Ensayo Cometa
Es un método sensible, rápido y relativamente de bajo costo para cuantificar daño en el ADN de células individuales. Las células con mayor daño del ADN muestran una migración aumentada desde el núcleo hacia el ánodo, generando una imagen similar a la cola de un cometa. La presencia de daño en el ADN se cuantifica midiendo la longitud de la cola o el “momento de la cola”. La importancia de este ensayo radica en la posibilidad de poder detectar fragmentación de simple o doble cadena del ADN espermático en una sola célula.
Ensayo de la Estructura Cromatínica del Espermatozoide (SCSA)
Es una técnica que desnaturaliza la molécula de ADN mediante una solución ácida o un tratamiento con calor y una vez desnaturalizada se tiñe con naranja de acridina (AO). El AO emite en verde cuando se intercala en cadenas de ADN dobles (ADN intacto), y en rojo si son de cadena sencilla (ADN fragmentado). El grado de daño de ADN se mide de forma cuantitativa por citometría de flujo y se expresa como DFI (DNA Fragmentation Index). Valores de DFI >27% están asociados con fallo de embarazo en TRA.
Dispersión de la Cromatina Espermática (SCD)
Esta técnica consiste en producir una descondensación diferencial de la cromatina en aquellos espermatozoides que tienen su ADN fragmentado respecto de aquellos que no lo tienen. Los espermatozoides con fragmentación no generan un halo de dispersión de la cromatina. Por el contrario, los que no están fragmentados dan lugar a grandes halos de dispersión. Se valora el tamaño de los halos de dispersión mediante microscopía de campo claro o de fluorescencia. Cabezas de espermatozoides con halos grandes o medianos significan que no hay daño en el ADN espermático, mientras que aquellos con halos pequeños o sin ellos sugieren un daño en el ADN.
Factores que Afectan la Descondensación y la Integridad del ADN
El estudio de la descondensación de la cromatina espermática también sirve como bioindicador del efecto de disruptores endócrinos. En un estudio, fue posible caracterizar el sistema de descondensación in vitro en el modelo de ratón al incubar los espermatozoides con distintos glicosaminoglicanos y GSH, determinándose las moléculas candidatas a ser posibles agentes descondensantes, las concentraciones y tiempos óptimos de incubación.
Se detectó un efecto sinérgico existente entre la heparina (utilizada como equivalente molecular del heparán sulfato) y el dermatán sulfato, los únicos glicosaminoglicanos estudiados con potencial capacidad descondensante. El efecto de estos dos glicosaminoglicanos también fue evaluado mediante la técnica de microscopia electrónica de transmisión, la cual puso en evidencia los cambios sufridos por los espermatozoides durante el proceso de descondensación.
Una vez caracterizado el sistema, se logró evidenciar el aumento de la descondensación in vitro que ocurre cuando se incuban los espermatozoides con endosulfán o etanol, como también luego de la ingesta de etanol. Además, fue posible evaluar el efecto del plaguicida sobre la fragmentación del ADN, así como también si existe alguna correlación entre la misma y la descondensación de la cromatina. Los efectos del etanol observados in vitro fueron confirmados en experimentos in vivo, en los cuales se intoxicaron ratones macho para realizar experimentos de fecundación in vitro. Estos resultados sugieren que el ensayo de descondensación espermática podría utilizarse como centinela para evaluar posibles efectos de tóxicos ambientales o ingeridos.
Estudios Comparativos y Parámetros Seminales
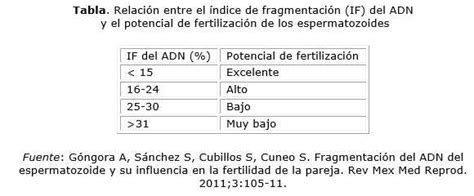
Un estudio comparativo en el Centro de Reproducción Asistida de Puebla, México (CREAFAM), aplicó una prueba de dispersión de ADN a muestras de semen de pacientes, clasificándolos en dos grupos: 76 pacientes con mala integridad espermática (fragmentación del ADN > 30%, grupo 1) y 77 con buena integridad espermática (fragmentación del ADN < 30%, grupo 2). Las características demográficas de ambos grupos fueron similares. Los pacientes del grupo 1 mostraron alteraciones espermáticas más severas. El porcentaje de varones según su diagnóstico seminal para el grupo de mala integridad fue mayor en casos de astenoteratozoospermia y normozoospermia.
La concentración de leucocitos en el semen no mostró diferencias significativas entre los grupos (p = 0.3529). En el grupo de buena integridad se registró un 5.93 ± 3.32% de fragmentación del ADN, mientras que en el grupo de mala integridad fue de 37.47 ± 40.65% (p < 0.0001). Los pacientes con mala integridad tuvieron alteraciones espermáticas más severas.
En el grupo de buena integridad, se observó una mayor proporción de espermatozoides con movilidad progresiva (50.56 ± 18.67% vs 25.12 ± 17.15%, p < 0.0001) y vitalidad (72.82 ± 13.92% vs 46.01 ± 23.36%, p < 0.0001) respecto del grupo de mala integridad. La concentración espermática fue de 57.07 ± 45.42 x 106/mL en el grupo de buena integridad y de 28.52 ± 29.83 x 106/mL en el grupo de mala integridad (p = 0.0063).
En cuanto a la morfología normal, el grupo de buena integridad presentó un 5.93 ± 3.32% frente a 4.24 ± 2.04% en el grupo de mala integridad (p = 0.0063). Se observó una menor presencia de leucocitos en el grupo de buena integridad (5.93 ± 3.32 vs 4.24 ± 2.04, p = 0.0063).
El estudio reveló que los varones con alta fragmentación del ADN espermático presentaron alteraciones espermáticas más severas. Esto incluye infertilidad masculina severa y factor tubárico.
tags: #proceso #de #descondensacion #de #cromatina #espermatozoides