Definición y Función General
El espermatozoide, también conocido como espermatozoón, es la célula sexual masculina (gameto) altamente especializada, diseñada para entregar el material genético del varón al óvulo durante la fertilización. Es una célula haploide, lo que significa que posee 23 cromosomas, la mitad de la dotación genética de las células somáticas. Su función principal es la formación de un cigoto totipotente al fusionar su núcleo con el del gameto femenino, un fenómeno que dará lugar, posteriormente, al embrión y al feto.
Además de perpetuar la especie por medio de la reproducción sexual, el espermatozoide también determina el sexo del futuro bebé. En función del reparto cromosómico que tiene lugar en la meiosis, la célula espermática poseerá el cromosoma X (sexo femenino) o el cromosoma Y (sexo masculino).
El espermatozoide fue descrito por primera vez en 1677 por el científico Anton van Leeuwenhoek, quien lo visualizó junto a su estudiante de medicina Johan Ham. Inicialmente, Ham pensó que se trataba de "animálculos" producto de la putrefacción del líquido seminal. Sin embargo, Leeuwenhoek supuso que eran un componente habitual del semen y realizó la primera descripción detallada. Posteriormente, Nicolás Hartsocker propuso en 1694 la teoría del homúnculo, creyendo que dentro de cada espermatozoide residía un ser humano en miniatura. Lazzaro Spallanzani, a finales del siglo XVIII, realizó experimentos pioneros de fecundación artificial en ranas y perros, demostrando el papel crucial del espermatozoide en la concepción.
Morfología General del Espermatozoide
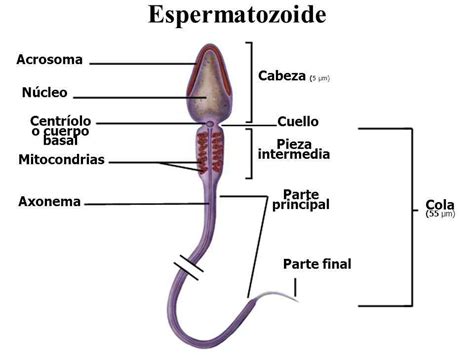
El espermatozoide es una célula alargada con una estructura distintiva que comprende una cabeza, un cuello y una cola, con una longitud total de unas 50-60 micras. La forma de los espermatozoides es similar en la mayoría de las especies, sobre todo en los mamíferos. Lo más peculiar del espermatozoide es que es la única célula humana con flagelo, la cola que le permite moverse.
Morfología Espermática Típica y Atípica
Por lo general, los espermatozoides tienen una cabeza lisa y ovalada con una larga cola. La morfología espermática se refiere al tamaño y la forma de los espermatozoides, siendo uno de los factores que los profesionales de atención médica analizan en un análisis de semen. En el laboratorio, los técnicos examinan el esperma al microscopio para ver si cumple unas normas estrictas.
Algunos espermatozoides tienen cabezas o colas que no son típicas. Esto puede incluir cabezas grandes o deformes, o colas torcidas o dobles. Tener muchos espermatozoides con estos problemas es algo común; en la mayoría de las muestras de semen, solo entre el 4 % y el 10 % de los espermatozoides cumplen normas estrictas. La mayoría de los expertos en fertilidad coinciden en que el papel de la morfología espermática en la predicción del embarazo no está claro, y no predice bien la falta de fertilidad. Tener semen que no cumpla las normas estrictas no significa que no se pueda tener hijos, aunque puede llevar más tiempo o requerir ayuda de un profesional de atención médica.
Partes del Espermatozoide
El espermatozoide se compone de tres partes principales: la cabeza, el cuello (o pieza intermedia) y la cola (o flagelo).
Cabeza
La cabeza del espermatozoide, de forma ovalada, tiene un tamaño entre 5 y 8 micras y es crucial para la fertilización. En ella se pueden diferenciar las siguientes partes:
- Núcleo: Es donde permanecen condensados los 23 cromosomas, es decir, la mitad de la información genética del futuro embrión. Esta parte es la única que entra dentro del óvulo y, por ello, es la más importante del espermatozoide. Su función es fusionarse con el núcleo del óvulo para completar la dotación genética del nuevo ser. La cromatina dentro del núcleo está altamente empaquetada, entre 5 y 10 veces más compactada que en cualquier otra célula, para proteger el ADN durante su transporte.
- Acrosoma: Es una estructura en forma de capucha que cubre los dos tercios anteriores del núcleo, desarrollándose a partir de la vesícula proacrosomal. Contiene enzimas proteolíticas como la hialuronidasa, acrosina y neuraminidasa, fundamentales para deshacer la zona pelúcida del óvulo y el cúmulo de células (cumulus oophorus) que lo rodea, permitiendo al espermatozoide penetrar en su interior.
- Membrana Plasmática: Rodea al acrosoma y al núcleo, separándolos del resto del cuerpo del espermatozoide. En su interior se encuentra una pequeña cantidad de citoplasma con altos niveles de ácidos grasos poliinsaturados.
Cuello o Pieza Intermedia
El cuello, o pieza intermedia, se sitúa entre la cabeza y la cola. Mide entre 6 y 12 micras, siendo ligeramente más larga que la cabeza y apenas visible al microscopio. Es un poco más gruesa que las demás partes del flagelo y puede contener residuos citoplasmáticos de la espermátida.
En su interior existen dos centriolos: el distal, que origina la pieza media, y el proximal, que se sitúa en la parte basal del núcleo y desaparece después de haber dado origen al flagelo. El cuello también contiene una placa basal de material denso que lo separa de la cabeza y donde se anclan 9 columnas proteicas (centriolos modificados) que continúan por toda la cola.
La pieza media (de unos 4 o 5 μm de longitud) posee una gran cantidad de mitocondrias concentradas en una vaina helicoidal, que proveen la energía necesaria (en forma de ATP) para el movimiento flagelar y el avance del espermatozoide. Estas mitocondrias se agrupan alrededor del axonema en su parte cercana al núcleo.
Cola o Flagelo
La cola del espermatozoide, también llamada flagelo, es una estructura larga, de aproximadamente 50 µm de longitud, cuya función principal es permitir la movilidad espermática mediante su movimiento ondeante o serpenteante. Este movimiento permite una velocidad de nado de aproximadamente 3 milímetros por minuto.
La cola se divide en tres partes: la pieza de conexión que une la cola con la cabeza, la pieza principal (la sección más larga y responsable del movimiento propulsor) y la pieza terminal (la región final, delgada y ahusada del flagelo). El axonema, eje funcional del flagelo, está compuesto por microtúbulos, moléculas chaperonas, proteínas fijadoras de calcio y proteínas quinasas/fosfatasas. Es una estructura de 9x2 pares de microtúbulos que corre a lo largo del flagelo. En la pieza media está rodeado por mitocondrias y capas de fibra densa; en la pieza principal, por una capa fibrosa; y en la final, está en contacto directo con la membrana plasmática. El movimiento del flagelo se produce por la activación de los complejos de ensamble y regulación de la dineína, cuya acción es impulsada por el ATP.
Cualquier alteración en la cola espermática que impida el movimiento progresivo de los espermatozoides será motivo de infertilidad masculina. Una vez que el espermatozoide llega hasta el óvulo y su núcleo entra en el interior, la cola se pierde.
Espermatogénesis: Formación del Espermatozoide
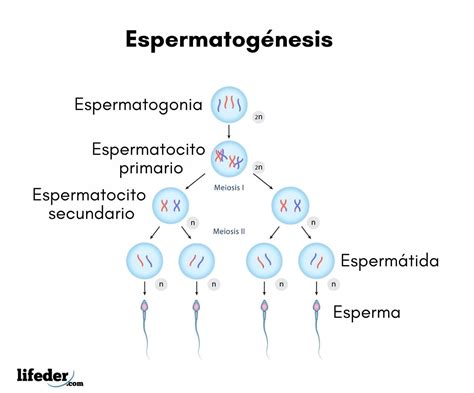
La espermatogénesis es el proceso continuo por el que las espermatogonias (células germinales primitivas) se transforman en espermatozoides maduros. Este proceso se produce en el interior del testículo, específicamente en los túbulos seminíferos, y se inicia en el periodo prepuberal del niño, entre los 11 y 15 años, manteniéndose durante prácticamente toda la vida del varón. La duración de todo el proceso de espermatogénesis es de aproximadamente 74 días (desde una espermatogonia hasta el espermatozoide final).
Fases de la Espermatogénesis
La espermatogénesis consta de tres fases principales:
- Fase Proliferativa: Las espermatogonias (células diploides con 46 cromosomas) se dividen por mitosis para mantener su "stock" y, a la vez, diferenciarse en espermatocitos primarios. A partir de cada espermatogonia, se producen cuatro espermatocitos haploides.
- Fase Meiótica: Durante esta fase, los espermatocitos primarios experimentan la primera división meiótica, formando los espermatocitos secundarios. Estos, a través de la segunda división meiótica, dan lugar a las espermátidas (células haploides y redondas con 23 cromosomas). Esta fase, que conlleva el apareamiento de los cromosomas con entrecruzamiento e intercambio de material genético, dura aproximadamente 24 días en humanos, de los cuales 23 días corresponden al estadio de espermatocito primario y solo 1 día a la segunda división meiótica.
- Espermiogénesis: En esta fase, las espermátidas se transforman en los espermatozoides finales mediante profundos cambios morfológicos. El citoplasma se desplaza hacia la cola y se elimina junto con los orgánulos y componentes innecesarios (formando el llamado cuerpo residual o gota citoplasmática). Se produce la formación del acrosoma (o gorro con alto contenido en glicoproteínas y enzimas proteolíticas), el desarrollo de la cola o flagelo, la transformación y compactación del núcleo en posición periférica. En el ser humano, este proceso dura unos 23 días.
Cuando termina la espermiogénesis, solo queda el proceso de espermiación, es decir, la liberación de los espermatozoides desde el epitelio seminífero hasta la luz de los túbulos.
Células de Sertoli
Las células de Sertoli son células somáticas que poseen abundante citoplasma, son ricas en elementos del citoesqueleto y vesículas. Son células muy plásticas, ya que contactan con la pared del túbulo seminífero, con células de Sertoli vecinas y con las células germinales.
Además, actúan como grandes recicladoras, eliminando las células que no completan la espermatogénesis y los cuerpos residuales. También se encargan de la translocación de las células que inician la meiosis hacia zonas más internas del túbulo, haciéndolas pasar al otro lado de la barrera hematotesticular y migrar hacia la luz del túbulo. Son esenciales para el soporte y nutrición de las células germinales en desarrollo.
Maduración, Movimiento y Fertilización
Maduración Post-Espermiogénesis y Transporte
Una vez formados en los túbulos seminíferos, los espermatozoides inmaduros pasan al epidídimo por acción de los elementos contráctiles de los túbulos. En el epidídimo, adquieren la capacidad de mover el flagelo. Durante la eyaculación, los espermatozoides se mezclan con los líquidos producidos por las vesículas seminales y la próstata para formar el semen. Estos líquidos son imprescindibles para la alimentación y supervivencia de los espermatozoides hasta alcanzar el óvulo.
La eyaculación es el reflejo de expulsión de los espermatozoides y el plasma seminal fuera del tracto reproductivo, resultado de la estimulación sensorial en el glande que causa contracciones musculares coordinadas. El eyaculado también contiene las secreciones de las glándulas anexas (vesículas seminales, próstata, glándulas bulbouretrales).
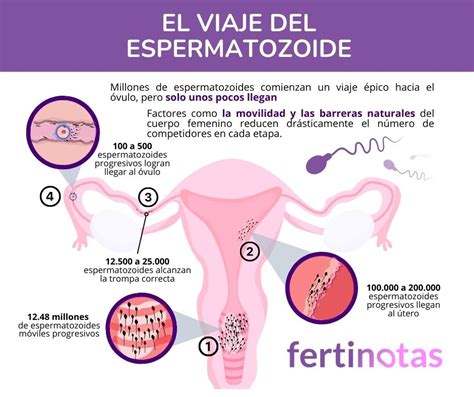
El Trayecto hasta el Óvulo
El movimiento del flagelo comienza después de la eyaculación, un proceso conocido como la activación del esperma. El movimiento es un bateo simétrico de la cola que permite el desplazamiento progresivo. El esperma pasa rápidamente a través del cuello y el útero femenino; a 10 minutos de la deposición del semen, ya se encuentran espermatozoides en la unión útero-tubal.
Desde allí, los espermatozoides pasan al reservorio del oviducto (istmo), caracterizado por cilios luminales epiteliales y plegamientos de la mucosa que forman criptas. En estas criptas, los espermatozoides son retenidos y pierden los factores decapacitantes (mucopolisacáridos y proteínas) aportados por las glándulas anexas. Este es el comienzo del proceso conocido como capacitación, donde el espermatozoide adquiere el potencial para hiperactivarse y para lograr la reacción acrosomal.
Cuando el espermatozoide alcanza el istmo del oviducto, inicia un movimiento asimétrico, amplio y acelerado del flagelo, característico de la hiperactivación. Este movimiento lo ayuda a liberarse de las criptas oviductales para avanzar a través del lumen y alcanzar el ámpula, atravesar el cúmulus ooforus (células de la granulosa que rodean el oocito) y aponerse a la zona pelúcida, donde es reconocido (reconocimiento entre gametos).
Reacción Acrosomal y Fecundación
La reacción acrosomal es un proceso de fusión de la membrana citoplasmática externa con la membrana acrosomal externa en la zona apical de la cabeza, liberando enzimas almacenadas dentro de esta vesícula exocítica. Esto es fundamental para el paso a través de la zona pelúcida.
Una vez que el espermatozoide alcanza el espacio perivitelino, se produce la adherencia entre la membrana plasmática de la zona ecuatorial espermática y las microvellosidades de la membrana citoplásmica del oocito. Luego, se fusionan ambas membranas, y de esta manera, el núcleo y demás organelas de la célula espermática ingresan al ooplasma del oocito.
Los depósitos de Ca++ almacenados entre la teca perinuclear y el núcleo del espermatozoide son liberados al ooplasma, junto con la "oscilina" (factor espermático). Este factor induce las oscilaciones de calcio que conducen a la activación del oocito. Esta activación comprende: oscilaciones intracelulares de Ca++, la expulsión del segundo cuerpo polar, la formación del pronúcleo femenino, el reemplazo de las protaminas por histonas en el núcleo espermático, la formación del pronúcleo masculino y la ubicación del centriolo espermático para formar el aster masculino necesario para la migración de su pronúcleo. Las oscilaciones de Ca++ también inducen la exocitosis del contenido enzimático de los gránulos corticales del oocito, modificando la zona pelúcida y la membrana del oocito para prevenir la poliespermia (entrada de múltiples espermatozoides).
Aspectos Moleculares y Epigenéticos del Espermatozoide
Regulación de la Movilidad Flagelar
La movilidad del esperma se desencadena por cambios en el medio iónico extracelular, la interacción con ligandos específicos y la glucosa, presentes en el líquido seminal y en el tracto reproductivo femenino. Estos cambios inducen señales citosólicas flagelares a través de la fosforilación de proteínas, canales de Ca++ y vías dependientes de nucleótidos cíclicos (GMPc y AMPc).
Los ligandos específicos incluyen progesterona y el esteroide sulfatado SAAF (Sperm Activating and Attracting Factor), que inducen la entrada de Ca++; el péptido activador de espermatozoide (PAS) y el péptido atrial natriurético (PAN), que actúan mediante un receptor de membrana o activación directa de la guanilil ciclasa ligada a membrana (GCm). Otros factores incluyen los de tipo olfatorio y odorante (hOR17-4) producidos por el oocito para inducir la quimiotaxis. Los cationes (Ca++, Na+, K+, H+) y aniones también modulan la movilidad espermática.
La activación de la movilidad flagelar implica la activación de ciclasas, un aumento transitorio de GMPc y AMPc, y la activación de la guanilil ciclasa transmembranal o soluble. Estos eventos conducen a la hiperpolarización de la membrana espermática, la alcalinización del citosol y la activación de las dineínas. La fosforilación y defosforilación de tirosinas, mediadas por la proteína quinasa dependiente de AMPc (PKA), son pasos cruciales. La PKA, con una estructura única en el espermatozoide, se encuentra anclada a los microtúbulos y cerca del brazo de dineína, explicando la rápida fosforilación de polipéptidos de la cadena liviana de este brazo.
¿Cómo se forman los espermatozoides?: Espermatogénesis
Remodelación de la Cromatina
La cromatina del esperma de los mamíferos es única, altamente organizada, condensada y compactada. El material genético del espermatozoide se encuentra de 5 a 10 veces más compactado que el de cualquier otra célula del cuerpo. Las protaminas condensan las cadenas de ADN y forman una unidad de empaquetamiento básica de la cromatina llamada toroide. Confieren un nivel mayor de empaquetamiento del ADN que el de las células somáticas, protegiendo la cromatina durante el transporte a través del tracto reproductivo masculino y femenino. Además, las protaminas son necesarias para el silenciamiento del genoma paterno y la reprogramación del patrón de impronta del gameto.
En la espermatogénesis, las histonas somáticas se reemplazan progresivamente por variantes de histonas específicas de los testículos. Durante la espermiogénesis, estas variantes son sustituidas por proteínas de transición (TP1 y TP2), un proceso que requiere la remodelación del ADN. Las proteínas de transición son necesarias para la normal condensación de la cromatina, reducir las roturas del ADN y prevenir defectos secundarios. Finalmente, en la elongación de las espermátides, las proteínas de transición se sustituyen por protaminas. En humanos, la relación P1/P2 es aproximadamente de 1.0, y las alteraciones en este cociente se asocian con infertilidad.
Las protaminas, con aproximadamente la mitad del tamaño de las histonas, son proteínas nucleares básicas ricas en argininas y residuos de cisteínas. Los altos niveles de arginina causan una carga neta positiva, facilitando su unión al ADN. Los residuos de cisteína facilitan la formación de múltiples puentes disulfuro (inter e antropotomías), esenciales para el empaquetamiento de orden superior de la cromatina.
Patrones Epigenéticos
El desarrollo de las células germinales primordiales hasta espermatozoides maduros es una etapa clave para la reprogramación epigenética. Estudios recientes muestran que las células germinales masculinas poseen un patrón único de metilación. Los patrones de metilación de promotores en el esperma, como la hipometilación, permitirían la expresión de genes específicos de las células germinales involucrados en la espermatogénesis; mientras que la hipermetilación daría lugar a la represión de la pluripotencia y de genes específicos de tejidos somáticos. La adquisición inicial de patrones epigenéticos se relaciona con la expresión de Dnmt3a y Dnmt3L, consistente con el papel de las enzimas DNMT3 como metiltransferasas de novo.
Alteraciones de los Espermatozoides y Patologías Asociadas
Existen varias alteraciones relacionadas con los espermatozoides que pueden dificultar la concepción de manera natural, provocando infertilidad masculina. Las anomalías más frecuentes incluyen:
- Oligozoospermia: Baja concentración de espermatozoides en el semen.
- Astenozoospermia: Alteración de la movilidad espermática debido a defectos que impiden o dificultan el avance de los espermatozoides. La movilidad Tipo A corresponde a espermatozoides con movimiento rectilíneo a una velocidad mayor de 25 micras/s (poco común, alrededor del 1%). El Tipo B se desplaza entre 5 y 24 micras/s, el Tipo C tiene una velocidad inferior a 5 micras/s (apenas se desplazan), y el Tipo D carece de movilidad.
- Teratozoospermia: Es cuando los espermatozoides presentan una mala morfología, ya sea en la cabeza, el cuello o la cola.
- Fragmentación del ADN: El material genético condensado en el núcleo de la cabeza espermática presenta roturas.
La consecuencia de estas alteraciones es la incapacidad de recorrer el camino para llegar al óvulo y/o problemas para la fecundación. Las alteraciones en la estructura del espermatozoide suelen tener su origen en errores durante la producción en los testículos o alteraciones adquiridas a lo largo de la maduración y recorrido. En casos de gravedad, puede ser necesario recurrir a técnicas de reproducción asistida.
Un epitelio seminífero sano presenta unas 4 o 5 capas de células germinales. Si se observan pocas capas, puede ser síntoma de que la espermatogénesis no es eficiente, o que existe una descamación de células que caen en la luz del túbulo, perdiendo su capacidad de proliferación. Un tejido testicular mal estructurado se relaciona con infertilidad (oligozoospermia). Otra patología conocida como síndrome de Sertoli-only se basa en la presencia única y exclusiva de células de Sertoli, las cuales colonizan la totalidad del túbulo. Normalmente, en pacientes con meiosis alterada, también se presentan alteraciones en el seminograma.
Factores Ambientales y Supervivencia
La producción diaria de células espermáticas implica que factores externos que afecten al varón en un momento puntual pueden interferir en el proceso de maduración. Por ello, se recomienda confirmar el diagnóstico de un factor masculino con un segundo estudio seminal tras uno o dos meses. La exposición a contaminantes ambientales (pesticidas, fertilizantes, disolventes), el consumo de tóxicos o el estrés pueden tener un efecto negativo en la calidad seminal.
Desde la eyaculación, los espermatozoides suelen vivir durante 2-5 días en el interior del aparato reproductor femenino, donde las condiciones de humedad y temperatura son idóneas. Sin embargo, si la eyaculación se produce en el exterior, la vida media de los espermatozoides se reduce drásticamente. La mayor capacidad fecundante se concentra durante las primeras 24 a 48 horas posteriores a la eyaculación.
Estudio de la Meiosis en el Varón
El estudio de la meiosis en el hombre adulto se puede realizar a nivel citogenético o histológico a partir del análisis del tejido testicular obtenido mediante biopsia. Esta prueba complementaria se solicita para orientar a la pareja sobre el proceso de fecundación in vitro más adecuado.
Análisis Citogenético
En el análisis citogenético, se obtienen y analizan las células precursoras de los espermatozoides en las diferentes fases de la meiosis de este tejido, destacando la profase I, la metafase I y la metafase II. Este estudio permite conocer si el reparto de los cromosomas se desarrolla correctamente. En caso de desarrollarse erróneamente, se obtendrán espermatozoides con un número anómalo de cromosomas: pueden ser diploides (doble dotación cromosómica) o aneuploides (un cromosoma de más o de menos).
Análisis Histológico
El estudio histológico requiere que los cortes sean completamente transversales y que la tinción esté realizada correctamente para diferenciar morfológicamente cada una de las células que forman parte del túbulo. Un epitelio sano muestra 4 o 5 capas de células germinales, distinguiéndose los núcleos de las espermatogonias en la base y las células más cercanas a la luz del túbulo a medida que avanza la meiosis.
tags: #quines #son #parts #espermatozoide