Las espermatogonias fetales representan las células germinales primordiales masculinas, cuyo desarrollo y migración inician el complejo proceso que culminará en la producción continua de espermatozoides en la vida adulta. Comprender su origen y las etapas tempranas de su desarrollo es fundamental para entender la espermatogénesis en su totalidad.
El Origen de las Células Germinales Masculinas Fetales
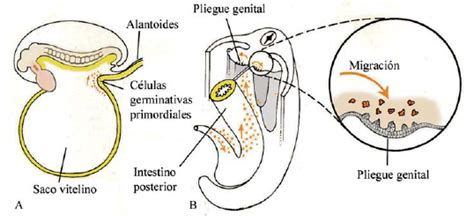
La formación de las espermatogonias comienza aproximadamente alrededor del día 24 del desarrollo embrionario en la capa endodérmica del saco vitelino. En esta etapa inicial, se producen unas 100 células germinales que inician un proceso de migración hacia los esbozos de los órganos genitales del embrión.
Hacia la cuarta semana de desarrollo, estas células germinales se acumulan en un número aproximado de 4000. En este punto crucial, el gen SRY juega un papel determinante al activar la formación de los testículos, lo que permitirá la posterior producción de espermatozoides. Sin embargo, este proceso de espermatogénesis activa no comenzará hasta la pubertad. En un corte histológico de un túbulo seminífero de un niño recién nacido, se observan túbulos con muy poca luz, junto con células de Sertoli inmaduras y algunas espermatogonias, que son las células madre del testículo. Esto confirma que la espermatogénesis, en su fase activa de producción, no se inicia hasta la pubertad.
Las Espermatogonias: Células Madre de la Línea Germinal
Las espermatogonias son las células germinales primordiales del macho, a partir de las cuales se producen los espermatozoides mediante mecanismos de mitosis y meiosis. Estas células se caracterizan por poseer un gran núcleo redondo u oval de cromatina condensada, en el que puede encontrarse el núcleo periférico y una vacuola nuclear.
Una vez llegada la edad reproductiva del hombre (pubertad o adolescencia), estas células se dividen múltiples veces para formar un tipo de célula denominada espermatocito primario. La espermatogénesis es el proceso continuo por el que las espermatogonias (células germinales primitivas) se transforman en espermatozoides, iniciando en el periodo prepuberal del niño, entre los 11 y 15 años. A diferencia de lo que ocurre en el sexo femenino, la formación del gameto masculino no comienza hasta la pubertad y luego dura toda la vida.
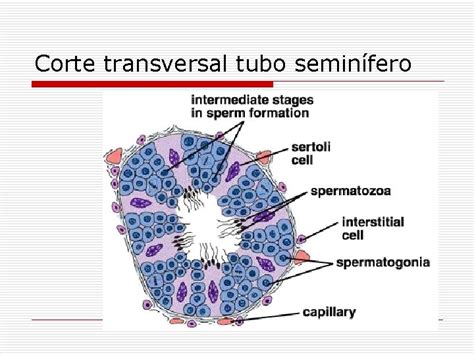
Las espermatogonias se encuentran en la base del túbulo seminífero y mantienen la reserva celular a través de su división. Se distinguen dos tipos principales:
- Espermatogonias tipo A: Siguen replicándose por mitosis, dando lugar a más espermatogonias de tipo A y a espermatogonias de tipo B. Estas células se caracterizan por poseer un gran núcleo redondo u oval de cromatina condensada, en el que puede encontrarse el núcleo periférico y una vacuola nuclear.
- Espermatogonias tipo B: Se dividen por mitosis produciendo más células de su tipo y maduran en grupos de aproximadamente 100 células, dando lugar a espermatocitos primarios. Estas se caracterizan por carecer de una vacuola nuclear.
En resumen, el proceso de la espermatogénesis consiste en el paso de una célula germinal, con 23 parejas de cromosomas (diploide), las espermatogonias, a convertirse en una célula con 23 cromosomas (haploide), los espermatozoides.
La Espermatogénesis: Un Proceso Continuo
La espermatogénesis es el proceso de diferenciación celular que conduce a la producción de espermatozoides. Se realiza en el interior de los testículos, en concreto en los túbulos seminíferos, pequeños conductos enrollados de 0,2 milímetros (mm) de diámetro. Los túbulos seminíferos están formados por una asociación de células germinales (espermatogonias, espermatocitos primarios y secundarios, espermátidas y espermatozoides) y células somáticas, llamadas células de Sertoli.
Las células de Sertoli son componentes de gran importancia en la regulación de la espermatogénesis. Forman la barrera hematotesticular, se ocupan de nutrir a los espermatozoides en desarrollo, favorecer su desarrollo y maduración, translocar las células que inician la meiosis hacia zonas más internas del túbulo, y eliminar las células que no completan la espermatogénesis y los cuerpos residuales. La capa basal de las células germinales se adosa a la membrana basal, rodeada por una lámina propia en la que existen varias capas de miofibroblastos y fibroblastos fusiformes.
Se sabe que para la producción y maduración de las células espermatogénicas es esencial que existan concentraciones elevadas de hormonas androgénicas, secretadas por las células de Leydig o intersticiales del mesotelio reticular. Las células de Sertoli secretan una proteína captadora de andrógenos que transporta la testosterona y la dihidrotestosterona hacia la luz del túbulo seminífero.
La espermatogénesis, en la especie humana, se produce en ondas a lo largo de los túbulos seminíferos, por lo que zonas adyacentes del mismo túbulo muestran espermatocitogénesis y espermiogénesis en diversas fases. Un ciclo espermatogénico tiene una duración aproximada de 2 meses (alrededor de 64 a 74 días en total).
La actividad enzimática de las polimerasas que participan en la espermatogénesis es óptima a una temperatura de aproximadamente 34 °C. Esta temperatura, ligeramente inferior a la corporal (1-3 °C), se mantiene gracias a la localización del testículo en la bolsa escrotal, que no contiene grasa y cuyos músculos reaccionan a la temperatura extendiéndose o contrayéndose.
Históricamente, los espermatozoides fueron observados por primera vez en el semen por Anton van Leeuwenhoek en 1677. Sin embargo, no fue hasta la llegada de la teoría celular y el descubrimiento de la naturaleza celular de los espermatozoides que se comenzó a estudiar de forma eficaz el desarrollo de las células reproductivas masculinas. Enrico Sertoli, en 1865, realizó el descubrimiento más importante en este campo con la demostración de las "células de soporte" en los túbulos seminíferos que llevan su nombre.
Fases de la Espermatogénesis
El proceso de espermatogénesis consta básicamente de tres fases principales:
- Fase Proliferativa (Espermatogónica): Las espermatogonias (diploides, con 46 cromosomas) se dividen por mitosis para mantener su stock y, a la vez, diferenciarse en espermatocitos primarios. Esta fase se caracteriza por la multitud de mitosis que producen muchas células precursoras de espermatozoides.
- Fase Meiótica (Espermatocitogénesis): Durante esta fase, los espermatocitos primarios experimentan la primera división meiótica, reduciendo la información genética a la mitad y formando espermatocitos secundarios haploides. Estos, a través de la segunda división meiótica, dan lugar a las espermátidas (células haploides y redondas). En total, de cada espermatocito primario (diploide) se obtienen cuatro espermátidas (haploides). Esta fase dura unos 24 días, de los cuales 23 corresponden al estadio de espermatocito primario.
- Espermiogénesis: Es la última etapa de la espermatogénesis, donde las espermátidas se transforman en espermatozoides maduros mediante una serie de cambios morfológicos y de diferenciación celular que duran unos 23-24 días.
Condensación del Núcleo
El núcleo se hace pequeño, denso y adquiere una forma aplastada, compactándose mucho más al cambiar las histonas por protaminas. De esta forma, no puede haber ni replicación ni transcripción (Fase G0 del ciclo celular). La inactividad transcripcional del núcleo hace que el espermatozoide sea dependiente de modificaciones postranscripcionales, como la fosforilación de proteínas.
Formación del Acrosoma
La vesícula acrosomal, que se localiza en la parte apical del núcleo, se aplana formando una verdadera capucha sobre el núcleo. El acrosoma se posiciona en el extremo apical y aparece rodeado por dos membranas, una externa y otra interna. Es un lisosoma especializado que contiene principalmente hialuronidasa, una enzima fundamental para degradar el ácido hialurónico de las células de la granulosa, permitiendo al espermatozoide atravesar las capas celulares que rodean al ovocito. Los depósitos de gránulos en el acrosoma también son importantes.
Formación del Flagelo
El flagelo se desarrolla a partir del centriolo distal y constituye un cilindro de 9 microtúbulos dobles que rodea dos únicos centrales (estructura 9+2), lo que se conoce como axonema. Durante la rotación del núcleo, se dispone en la base de la cabeza. El flagelo completo consta de cuello, pieza media, pieza principal y pieza final, siendo fundamental para la movilidad espermática. Los centriolos del espermatozoide cumplen un papel esencial en la fecundación, ya que el ovocito carece de ellos; son necesarios para el posicionamiento de los cromosomas en la placa metafásica durante la primera división del cigoto.
Reducción del Citoplasma
Gran parte del citoplasma de la espermátida es fagocitado por las células de Sertoli o se abandona en el interior de los túbulos, formando la llamada gota citoplasmática. El espermatozoide maduro es una célula muy especializada con un tamaño reducido.
Regulación Hormonal
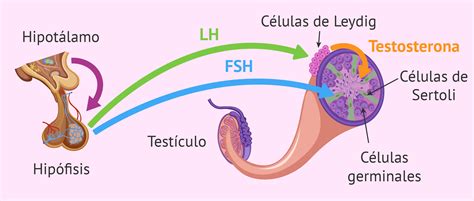
La espermatogénesis está regulada hormonalmente por el eje hipotálamo-hipofisario-gonadal mediante un mecanismo de retroalimentación negativa. Las hormonas implicadas son:
- GnRH (Hormona liberadora de gonadotropina).
- LH (Hormona luteinizante): Secretada por la hipófisis, activa la liberación de testosterona por las células de Leydig.
- FSH (Hormona folículoestimulante): Secretada por la hipófisis, actúa sobre las células de Sertoli para nutrir los espermatozoides y favorecer su desarrollo y maduración.
- Testosterona: Sintetizada por las células de Leydig en respuesta a la LH, activa genes que promueven la diferenciación de las espermatogonias.
- Inhibina: Liberada por las células de Sertoli, inhibe la liberación de FSH por la hipófisis, deteniendo así la espermatogénesis.
Morfología del Espermatozoide Maduro
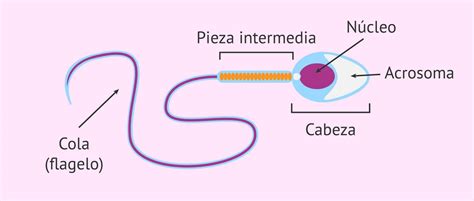
El espermatozoide es la célula reproductiva masculina especializada en transportar el material genético paterno para la fertilización del óvulo. Es una célula alargada con una estructura distintiva que comprende una cabeza, un cuello y una cola.
- La cabeza: De forma ovalada y tamaño entre 5 y 8 micras, contiene los cromosomas de la herencia y lleva en su parte anterior un pequeño saliente o acrosoma, cuya misión es perforar las envolturas del óvulo. El núcleo alberga los 23 cromosomas, con la cromatina altamente empaquetada. La membrana plasmática rodea el acrosoma y el núcleo, protegiéndolos.
- El cuello o pieza intermedia: Mide entre 6 y 12 micras, es un poco más larga que la cabeza y en ella se agrupan las mitocondrias alrededor del axonema en forma de hélice, formando la pieza media, la cual proporciona la energía necesaria para el movimiento.
- El flagelo o cola: Es una estructura larga, de aproximadamente 50 µm, fundamental para la movilidad espermática, permitiendo una velocidad de nado de alrededor de 3 milímetros por minuto. Está compuesto por las piezas media, principal y terminal.
Maduración y Movilidad del Espermatozoide
Una vez formados en los túbulos seminíferos, los espermatozoides inmaduros pasan al epidídimo. En su tránsito por el epidídimo, que dura alrededor de 10 días, adquieren la capacidad de mover el flagelo y la movilidad necesaria. Aquí se almacenan en su cola hasta el momento de la eyaculación, momento en el que son transportados a través de los conductos deferentes, mezclándose con el líquido seminal proveniente de las vesículas seminales y la próstata para formar el semen, y finalmente son expulsados por la uretra.
El movimiento del flagelo comienza después de la eyaculación, en un proceso conocido como la activación del esperma. El movimiento característico consiste en un bateo simétrico de la cola que hace que el espermatozoide se desplace de forma progresiva. El esperma pasa rápidamente a través del cuello y el útero de la hembra; a los 10 minutos de la deposición del semen en el tracto femenino, se encuentran espermatozoides en la unión útero-tubal. La movilidad del esperma se desencadena por cambios en el medio iónico extracelular, por interacción con ligandos específicos y por glucosa, presentes en el líquido seminal y en el tracto reproductivo femenino. Estos cambios inducen señales citosólicas flagelares, a través de la fosforilación de proteínas, de canales de Ca++ y de vías dependientes de nucleótidos cíclicos (GMPc y AMPc).
Desde la unión útero-tubal, los espermatozoides pasan a lo que se conoce como el reservorio del oviducto (istmo), caracterizado por cilios luminales epiteliales y plegamientos de la mucosa que forman criptas. En las criptas oviductales, los espermatozoides pierden los factores decapacitantes (mucopolisacáridos y proteínas aportados por las glándulas anexas). Este es el comienzo del proceso conocido como capacitación, que indica el potencial que adquiere el espermatozoide para hiperactivarse y para lograr la reacción acrosomal.
Cuando el espermatozoide alcanza el istmo del oviducto, inicia un movimiento asimétrico, amplio y acelerado del flagelo, característico de la hiperactivación. Este movimiento lo lleva a moverse en círculos y lo ayuda a liberarse de las criptas oviductales para avanzar a través del lumen y alcanzar el ámpula, atravesar el cúmulus ooforus (células de la granulosa que rodean el ovocito) y aponerse a la zona pelúcida donde es reconocido (reconocimiento entre gametos).
Fecundación y Determinación del Sexo Fetal
Para que se produzca un nuevo ser, es necesario que el óvulo y el espermatozoide se fusionen en un proceso llamado fecundación.
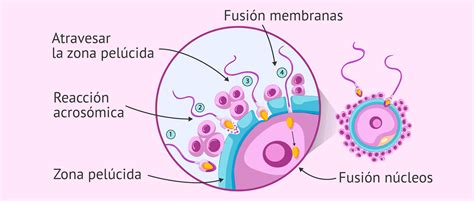
La reacción acrosomal es un proceso de fusión de la membrana citoplasmática externa con la membrana acrosomal externa en la zona apical de la cabeza del espermatozoide. De aquí se liberan enzimas que estaban almacenadas dentro de esta vesícula exocítica, lo cual es fundamental para el paso a través de la zona pelúcida. Una vez que el espermatozoide alcanza el espacio perivitelino, se produce la adherencia entre la membrana plasmática de la zona ecuatorial espermática y las microvellosidades de la membrana citoplásmica del ovocito. Luego, se fusionan las dos membranas, y de esta manera, el núcleo y demás orgánulos de la célula espermática ingresan al ooplasma del ovocito.
Los depósitos de Ca++ almacenados entre la teca perinuclear y el núcleo del espermatozoide son liberados al ooplasma, junto con la "oscilina" (factor espermático). A este factor se le atribuye la acción de inducir las oscilaciones de calcio que conducen a la activación del ovocito. Esta activación comprende: oscilaciones intracelulares de Ca++, la expulsión del segundo cuerpo polar, la formación del pronúcleo femenino, el reemplazo de las protaminas por histonas en el núcleo espermático, la formación del pronúcleo masculino y la ubicación del centriolo espermático para formar el aster masculino.
Cómo se produce la fecundación | Animación 3D
Gametogénesis y Cromosomas Sexuales
Las células humanas contienen 46 cromosomas cada una, a excepción de los gametos (espermatozoides y óvulos), que cuentan con solo 23 cromosomas cada uno para que al juntarse en la fecundación reúnan los 46 habituales y formen un cigoto. Los óvulos siempre tienen el cromosoma sexual X, puesto que las mujeres poseen dos cromosomas X (XX). Los espermatozoides, sin embargo, pueden tener un cromosoma X o un cromosoma Y. Por lo tanto, si es un espermatozoide con un cromosoma X el que fecunda al óvulo, el cigoto resultante será de sexo femenino (XX). Por el contrario, si el espermatozoide que fecunda al óvulo tiene un cromosoma Y, el cigoto será de sexo masculino (XY).
En definitiva, genéticamente, la probabilidad de tener un niño o una niña depende de si el espermatozoide que fecunda al óvulo es X o Y.
Factores que Influyen en la Probabilidad del Sexo
Aunque la determinación del sexo sea un proceso aleatorio, algunos estudios sugieren que en la probabilidad de tener un niño o una niña influye el día en el que tenga lugar el coito respecto a la ovulación. Esto se debe a que los espermatozoides con copia X e Y presentan algunas características diferentes:
- Espermatozoides Y: Son más rápidos, pero sobreviven menos tiempo en el tracto genital femenino.
- Espermatozoides X: Son más lentos, pero son más resistentes y, por tanto, sobreviven durante más tiempo.
Estas hipótesis sugieren que si el coito tiene lugar cuando ya se ha ovulado, los espermatozoides con copia Y alcanzarán antes el óvulo, aumentando las probabilidades de tener un niño. Por otro lado, si se produce antes de la ovulación, los espermatozoides con un cromosoma X pueden sobrevivir más horas hasta que el óvulo alcance las trompas de Falopio, incrementando las posibilidades de tener una niña. No obstante, no existe una garantía de que el bebé vaya a ser de un sexo u otro si se toman estas medidas, ya que son solo hipótesis.
Avances en la Determinación del Sexo Fetal
Hoy en día, gracias a los avances científicos, es posible conocer el sexo del bebé en las primeras semanas de gestación de forma certera y segura. A partir de la sexta semana de gestación, se pueden encontrar en el torrente sanguíneo de la madre fragmentos de ADN libre fetal que proceden de las células de la placenta. Para poder realizar la prueba con una concentración adecuada de ADN libre fetal, se recomienda esperar a la semana 10.
Este descubrimiento ha supuesto un gran avance, ya que permite saber si el bebé presenta alguna alteración cromosómica mediante una prueba de cribado no invasiva con una muestra de sangre de la madre. Además de detectar posibles alteraciones cromosómicas en el bebé, también se puede determinar el sexo fetal y, por tanto, ver si existen anomalías en los cromosomas sexuales.
El test de cribado analiza el ADN de las células de la placenta. La detección de un cromosoma Y es compatible con la presencia de un feto de sexo masculino. Por lo que, si no se detecta ningún fragmento de cromosoma Y, se infiere que el sexo es femenino. En los casos de embarazos gemelares, si hay presencia del cromosoma Y, puede determinarse que, al menos, uno de los fetos es masculino, y si hay ausencia de este, se puede asumir que ambos fetos son femeninos.
Este tipo de pruebas también pueden identificar trastornos de los cromosomas sexuales, que se dan cuando el número de cromosomas sexuales que tiene el bebé no es el habitual. Entre estas alteraciones, que pueden implicar problemas físicos, de aprendizaje e infertilidad, las más comunes son el síndrome de Turner (falta de un cromosoma X en mujeres), el síndrome de Klinefelter (dos cromosomas X y uno Y en varones), el síndrome Triple X (tres cromosomas X en mujeres) o el síndrome XYY (un cromosoma X y dos Y en varones).
tags: #mis #espermatogonias #fetales