La fecundación, la unión de los gametos masculino y femenino (espermatozoide y óvulo), es un evento fundamental para el inicio de una nueva vida. Este complejo proceso se caracteriza por una intrincada serie de interacciones moleculares y celulares, donde el calcio (Ca²⁺) emerge como un mensajero intracelular clave. La regulación de la concentración de calcio citosólico es vital, ya que está implicada en numerosos eventos celulares, desde el control del ciclo celular hasta la contracción muscular y la secreción, siendo un "interruptor" principal que activa tanto al espermatozoide como al ovocito.
Regulación del Calcio Intracelular: Un Mensajero Universal
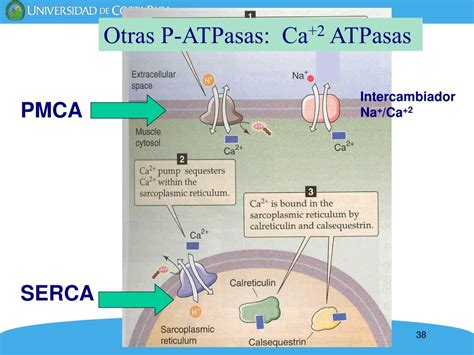
En las células eucariotas, la concentración de Ca²⁺ libre citosólico ([Ca²⁺]i) se mantiene en aproximadamente 100 nM, significativamente menor que la concentración de Ca²⁺ libre extracelular ([Ca²⁺]o), que suele ser de 1-2 mM. Este gradiente se sostiene mediante la extrusión activa de Ca²⁺ al medio extracelular a través de bombas de Ca²⁺, denominadas Ca²⁺-ATPasas. Entre ellas, la PMCA (plasma membrane Ca²⁺-ATPase), ubicada en la membrana plasmática, utiliza la energía de la hidrólisis de ATP para bombear Ca²⁺ fuera de la célula contra su gradiente de concentración.
Además, el retículo endoplasmático (RE) actúa como un reservorio intracelular de Ca²⁺, manteniendo concentraciones intraluminales muy superiores a las citosólicas. La SERCA (sarco(endo)plasmatic reticulum Ca²⁺-ATPase) es la bomba que transporta activamente Ca²⁺ desde el citosol hacia el interior del RE. Tanto en la membrana plasmática como en la del RE existen canales de Ca²⁺ que permiten la entrada o salida de este ion, facilitando incrementos transitorios de [Ca²⁺]i en respuesta a diversos estímulos, como la fusión espermatozoide-ovocito.
Estos incrementos deben ser contrarrestados para que los niveles de Ca²⁺ citosólicos retornen a su estado basal, asegurando que la señalización mediada por Ca²⁺ sea limitada y reversible.
Capacitación Espermática y Reacción Acrosomal
Para que los espermatozoides de mamíferos adquieran la capacidad de fertilizar un ovocito, deben someterse a un proceso crucial denominado capacitación durante su migración por el tracto reproductor femenino. Este proceso implica una serie de complejos cambios celulares y bioquímicos.
Cambios durante la Capacitación
- Reducción de Colesterol: Disminuye la rigidez de la membrana plasmática, facilitando re-arreglos de lípidos y proteínas.
- Entrada de Iones: Se observa un aumento en la entrada de iones como Ca²⁺ y HCO₃⁻.
- Activación de Vías de Señalización: Estos iones activan la adenilato ciclasa soluble ADCY10, lo que lleva a un aumento de AMPc y la activación de PKA. La entrada de HCO₃⁻ también contribuye a la hiperpolarización de la membrana y al aumento del pH intracelular (pHi).
Como resultado de la capacitación, el espermatozoide adquiere dos capacidades fisiológicas esenciales:
- Movilidad Hiperactivada: Un patrón de movimiento vigoroso que le permite penetrar medios viscosos y la zona pelúcida del óvulo.
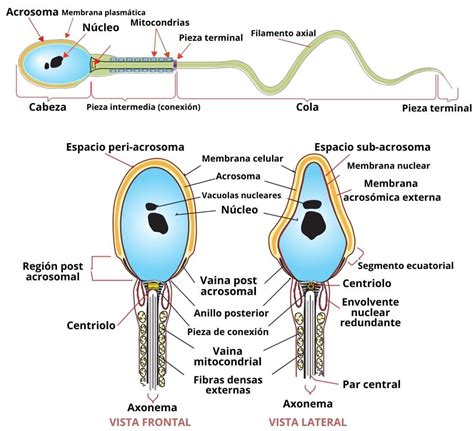
- Capacidad para la Reacción Acrosomal: La exocitosis acrosomal es un proceso especializado donde se fusionan la membrana citoplasmática y la membrana acrosomal externa en la cabeza del espermatozoide. Esto resulta en la liberación de enzimas hidrolíticas almacenadas en el acrosoma y la exposición de moléculas necesarias para la fusión con el ovocito.
Históricamente, ya a principios de 1950, se demostró que la reacción acrosomal es dependiente de calcio. La inducción de esta reacción puede lograrse in vitro mediante factores como el fluido folicular, la progesterona o ionóforos de calcio, lo que confirma el papel central del Ca²⁺ en este proceso.
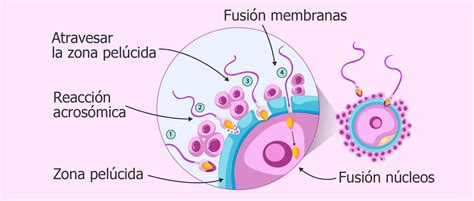
Etapas de la Fecundación y la Interacción Gamética
Para que ocurra la fecundación, los espermatozoides deben superar varias barreras y realizar interacciones específicas con el ovocito.
1. Penetración de la Corona Radiada
El proceso de fecundación se inicia con la penetración de los espermatozoides a través de la corona radiada, una capa de células que rodea al óvulo. Los espermatozoides consiguen atravesar esta capa gracias a la liberación de la enzima hialuronidasa y al movimiento de su flagelo.
2. Penetración de la Zona Pelúcida
Una vez superada la corona radiada, los espermatozoides se encuentran con una segunda barrera: la zona pelúcida, una membrana traslúcida que rodea directamente al óvulo. Para atravesar esta capa, la cabeza del espermatozoide establece contacto con el receptor ZP3 de la zona pelúcida del óvulo, lo que desencadena la reacción acrosómica. Esta reacción implica la liberación de enzimas hidrolíticas denominadas espermiolisinas, que disuelven la zona pelúcida para permitir el paso del espermatozoide.
Estudios recientes han identificado la importancia de la glicoproteína ZP2 en la zona pelúcida, ya que los espermatozoides se adhieren a ella antes de la fecundación, y su ausencia en ratones hembra provoca infertilidad.
3. Fusión de Membranas
Aunque se necesita más de un espermatozoide para degradar la zona pelúcida, finalmente solo uno logrará entrar en el óvulo. Cuando el espermatozoide entra en contacto con la membrana plasmática del óvulo, se desencadenan tres procesos cruciales en el gameto femenino:
- La formación del cono de fecundación, que permite la fusión de las membranas del óvulo y del espermatozoide, facilitando la entrada de la cabeza, la pieza intermedia y la cola del espermatozoide al citoplasma ovocitario.
- La despolarización instantánea de la membrana del óvulo.
- La liberación de gránulos corticales al espacio perivitelino.
Estos dos últimos mecanismos, conocidos como reacción cortical o de zona, son esenciales para evitar la polispermia (la entrada de múltiples espermatozoides), modificando la permeabilidad de la membrana del óvulo e impidiendo la penetración de otros espermatozoides.
La Ola de Calcio en el Ovocito: Activación Post-Fecundación
La ola de calcio, un aumento repentino y transitorio en la concentración de Ca²⁺ dentro del citoplasma del ovocito, es un fenómeno clave que se produce durante la fertilización y es fundamental para iniciar el desarrollo embrionario. Este incremento de [Ca²⁺]i puede alcanzar valores de 1-3 μM y se manifiesta como elevaciones transitorias y repetitivas, conocidas como ondas de calcio, las cuales se generan en un punto localizado del ovocito y se transmiten rápidamente al resto del citosol.
Rol de los Ovocitos en la Fecundación - LA PIZARRA A UN CLIC CIENCIAS BIOLOGÍA (PAES)
Inicio y Transmisión de las Ondas de Calcio
Las ondas de calcio son iniciadas por una actividad enzimática específica del espermatozoide. Tras la fusión de las membranas del espermatozoide y el ovocito, la enzima fosfolipasa C zeta (PLCζ) del espermatozoide es liberada al citosol del ovocito. La actividad de PLCζ hidroliza el lípido de membrana fosfatidilinositol 4,5-bisfosfato (PIP2), produciendo inositol 1,4,5-trisfosfato (InsP3) y 1,2-diacilglicerol (DAG).
- El InsP3, al ser soluble, difunde al citosol y se une a su receptor, InsP3R, localizado en la membrana del RE.
- El InsP3R, que funciona como un canal de Ca²⁺, se activa y permite la salida de Ca²⁺ desde el interior del RE hacia el citosol, induciendo la primera subida de [Ca²⁺]i en el ovocito.
Una vez liberado el Ca²⁺ del RE, el ovocito activa mecanismos para restaurar los niveles basales de Ca²⁺ citosólico. Esto incluye la actividad de las bombas PMCA (extrusión de Ca²⁺ al exterior) y SERCA (bombeo de Ca²⁺ al interior del RE), lo que permite que el nivel basal de Ca²⁺ citosólico se recupere en pocos segundos, preparando el terreno para una nueva onda.
La PLCζ permanece activa en el ovocito durante varias horas, hasta la formación de los pronúcleos. Su localización citosólica, en ausencia de membrana nuclear, asegura un tren continuo de ondas de Ca²⁺, permitiendo que el ovocito salga del bloqueo meiótico en el que se encuentra y complete su maduración (meiosis II).
Regulación por Depósitos y Canales SOCE en el Ovocito
Para garantizar que los depósitos intracelulares de Ca²⁺ en el RE no se agoten y la señalización mediada por Ca²⁺ se mantenga, existe un sistema de canales de Ca²⁺ en la membrana plasmática regulados por el nivel de Ca²⁺ intraluminal en el RE. Estos canales, denominados canales de Ca²⁺ regulados por depósitos intracelulares (canales SOC), permiten la entrada de Ca²⁺ desde el exterior celular al citosol, lo que a su vez permite que la SERCA bombee Ca²⁺ de nuevo al RE, restaurando así los niveles intraluminales. Este proceso se conoce como entrada de Ca²⁺ regulada por depósitos intracelulares (SOCE).
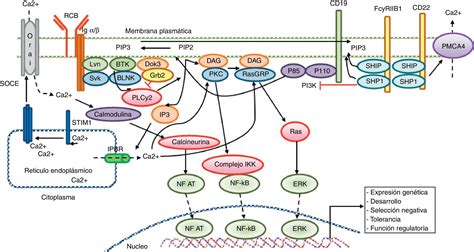
Papel de STIM1, ORAI1 y TRPC
Aunque los mecanismos exactos de cómo los canales SOC detectan el nivel de Ca²⁺ en el RE aún se están investigando, se sabe que la proteína STIM1 (stromal interaction protein 1) es fundamental. STIM1 es una proteína transmembranal del RE que actúa como sensor de Ca²⁺ intraluminal gracias a su dominio "EF-hand". Cuando la concentración de Ca²⁺ en el RE disminuye, STIM1 sufre oligomerización y se aproxima a la membrana plasmática, formando yuxtaposiciones RE-membrana plasmática y activando los canales SOC de la membrana plasmática. Así, STIM1 establece un puente para reponer el Ca²⁺ en el RE y asegurar la continuidad de las ondas de Ca²⁺.
Estudios recientes sugieren que la activación de los canales SOC es un proceso muy temprano en la fecundación, lo que implica que la vía SOCE no solo mantiene la señalización de Ca²⁺ a largo plazo, sino que también contribuye a la generación de las primeras ondas. Esto se apoya en el hecho de que la señalización de Ca²⁺ cesa inmediatamente en soluciones libres de Ca²⁺ y que la relocalización de STIM1 ocurre concomitantemente con la primera onda.
La identidad molecular de los canales SOC activos en el ovocito es crucial. En 2006, se describió ORAI1 (también conocida como CRACM1) como una proteína de membrana plasmática que constituye el canal responsable de la entrada de Ca²⁺ regulada por depósitos intracelulares. Además, STIM1 interactúa directamente con los canales de la familia TRPC (transient receptor potential canonical channels), incluyendo TRPC1, TRPC4 y TRPC5, e incluso forma interacciones heteroméricas con TRPC1 y ORAI1, dando lugar a complejos TRPC1-ORAI1-STIM1 que funcionan como canales SOC con diferentes propiedades de conductancia.
Implicaciones y Desafíos en Reproducción Asistida
La determinación de las proteínas que actúan como canales SOC en el ovocito es fundamental para el avance en la biología reproductiva y para aplicaciones prácticas en técnicas de reproducción asistida. Además, se ha constatado que la entrada de Ca²⁺ a través de estos canales es muy sensible a las especies reactivas de oxígeno (ROS), que pueden generarse en exceso en los medios de cultivo in vitro debido a la mayor tensión de oxígeno atmosférico. Esta desregulación inducida por el estrés oxidativo, a través de una sobrecarga de Ca²⁺ por los canales SOC descontrolados, puede afectar negativamente al ovocito.
Consecuencias de la Fusión de Gametos
Tras la entrada del espermatozoide, el ovocito se activa y termina su maduración (meiosis II), lo que implica la reducción del número de cromosomas. Se libera el segundo corpúsculo polar, y los cromosomas del ovocito forman el pronúcleo femenino. Por su parte, el núcleo del espermatozoide se hincha para formar el pronúcleo masculino.
Ambos pronúcleos, que contienen 23 cromosomas cada uno, se aproximan y sus membranas desaparecen para que sus cromosomas se fusionen. Este proceso restablece la dotación cromosómica normal del ser humano (46 cromosomas) y culmina con la formación del cigoto humano, la primera célula del nuevo organismo.
Determinación del Sexo
En este momento, también se establece el sexo del futuro bebé:
- Cigoto masculino: cromosomas sexuales XY.
- Cigoto femenino: cromosomas sexuales XX.
Dado que el óvulo siempre es portador del cromosoma X, el sexo del embrión está determinado por el espermatozoide, que puede ser portador de un cromosoma X o un cromosoma Y.
tags: #espermatozoide #entrada #calcio