El desarrollo embrionario es un proceso trascendental donde, a partir de un grupo indeterminado de células pluripotenciales, se establecen los esbozos de los futuros órganos en sus posiciones correctas y las células se comprometen a sus destinos de forma prácticamente irreversible. Aunque las etapas previas a la gastrulación difieren entre mamíferos y otros vertebrados debido a variaciones en la segmentación, una vez formado el disco embrionario y comenzada la gastrulación, los mecanismos de desarrollo para establecer el futuro organismo siguen patrones muy similares.
Este hecho relevante, denominado convergencia evolutiva, subraya la importancia del mecanismo de gastrulación, considerado un "cuello de botella" por el que han de pasar todos los vertebrados para la formación de un nuevo organismo. Por esta razón, gran parte de los datos más relevantes sobre la gastrulación se han obtenido del estudio de animales más accesibles para la experimentación, como los embriones de aves.
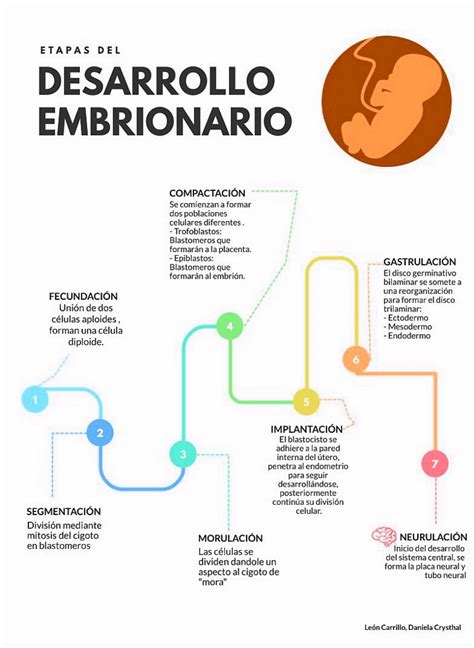
Fases Iniciales del Desarrollo Embrionario
Del Cigoto al Blastocisto
En un ciclo menstrual normal, un óvulo se desprende de uno de los ovarios aproximadamente 14 días después de la última menstruación, un proceso conocido como ovulación. Este óvulo entra en una de las trompas de Falopio. Durante la ovulación, el moco cervical se vuelve más fluido y elástico, facilitando la entrada de los espermatozoides. En aproximadamente 5 minutos, los espermatozoides se desplazan desde la vagina, a través del cuello uterino, hacia el interior del útero y hasta el extremo de una trompa de Falopio, donde habitualmente se produce la fecundación.
Si un espermatozoide penetra en el óvulo, este queda fecundado. Las células ciliadas que recubren la trompa de Falopio ayudan a arrastrar al óvulo fecundado (cigoto) hacia la cavidad uterina. Las células del cigoto se dividen repetidamente (se segmentan) mientras desciende por la trompa de Falopio. El cigoto tarda de 3 a 5 días en entrar en el útero. Una vez dentro, las células continúan dividiéndose y se convierten en una bola hueca denominada blastocisto, que se implanta en la pared del útero aproximadamente 6 días después de la fecundación.
En el caso de las técnicas de reproducción asistida (RA), el embrión es transferido a la cavidad uterina en la etapa de blastocisto o en alguna de las fases previas a su formación. La embriocinética, una de las nuevas tecnologías, aporta constantemente conocimientos sobre las particularidades de este proceso.
Formación del Disco Embrionario y la Gastrulación
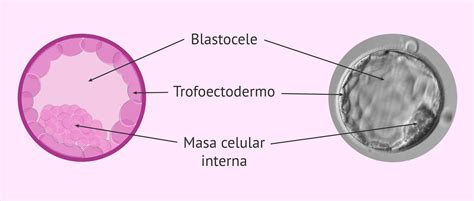
Simultáneamente con la implantación, la masa celular interna (MCI) del blastocisto experimenta una serie de cambios vertiginosos que conducen a la formación del embrión propiamente dicho. Aunque cuando el blastocisto es transferido la MCI está formada por un número relativamente pequeño de células (aproximadamente 50), la proliferación celular incrementa rápidamente este número sin apenas reducción del volumen celular, característica que marcó las divisiones anteriores durante la segmentación.
Al mismo tiempo que se produce este incremento, la MCI deja de ser una estructura sin organización aparente y, mediante procesos complejos, se constituye en una masa ligeramente aplanada o discoidal que recibe el nombre de disco embrionario. Los mecanismos mediante los cuales se establecen los ejes corporales en los vertebrados son muy complejos y, posiblemente, ya estén establecidos incluso desde la propia fecundación.
Gastrulación y Establecimiento de los Ejes Embrionarios
Formación de las Capas Germinativas
En fases precoces del desarrollo, las células del ectoblasto (epiblasto) inician dos procesos fundamentales y concomitantes: proliferación (división celular) y migración. Estos procesos celulares hacen que numerosas células del epiblasto se dirijan hacia el hipoblasto, desplazando sus células para ser sustituidas por una nueva capa celular, el endodermo.
El inicio de la gastrulación se caracteriza por los cambios morfogenéticos, ya que las células del ectodermo se dividen y migran para formar la tercera capa, el mesodermo. Las células ectodérmicas, con gran capacidad de proliferación, están sometidas a diferentes corrientes de migración celular, fundamentalmente en dos direcciones: látero-medial y rostro-caudal.
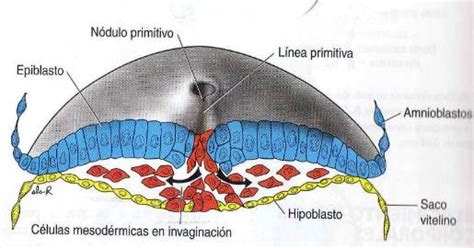
La Línea Primitiva y el Nódulo de Hensen
La línea primitiva es el aspecto morfológico que presentan las células al ingresar a través de esta estructura para formar el mesodermo. Las células que siguen una migración rostro-caudal se invaginan a nivel de la zona más rostral de la línea primitiva y forman sucesivamente, a lo largo de la capa media del embrión, una estructura alargada y central denominada notocorda o mesodermo axial, en el eje embrionario longitudinal.
Un análisis detallado de la migración celular a través de la línea primitiva permite distinguir un sector funcionalmente diferente en el extremo más rostral, donde se invaginan las células que darán lugar a la notocorda. Esta zona inicial, rostral, de la línea primitiva se denomina el organizador o, en ratones, el nódulo de Hensen. Es interesante destacar que la notocorda no llega a configurarse entre las capas ectodérmica y endodérmica en las posiciones más craneales o rostrales del embrión, debido a una íntima unión entre el ectodermo y el endodermo a ese nivel.
Desarrollo embrionario en animales
Desarrollo Progresivo Rostro-Caudal
Las características dinámicas de la gastrulación determinan el aspecto progresivo del desarrollo en estas fases iniciales. Dado que las células de las posiciones rostrales inician estos procesos de división y migración antes que las células de las posiciones embrionarias más caudales, el desarrollo embrionario sigue una secuencia rostro-caudal. Un embrión mostrará sus tres capas embrionarias (ecto-, meso- y endodermo) en el sector rostral (craneal o cefálico), mientras que el sector caudal aún estará en fase de formación de la línea primitiva.
Desarrollo y Diferenciación de las Capas Embrionarias
Crecimiento Tridimensional y Especialización
A partir de la gastrulación, el embrión comienza a crecer para adquirir progresivamente su aspecto tridimensional. Para ello, el embrión muestra dos vías fundamentales: cada una de las tres capas se desarrollará para formar los órganos y aparatos específicos, y el cuerpo embrionario se incurvará en sentido céfalo-caudal y lateral para configurar el cuerpo tridimensional. El desarrollo de cada capa es coincidente y concomitante con el de las otras dos.
Además, durante el proceso morfogenético de cada capa, existen importantes interacciones tisulares, celulares y moleculares entre los diferentes componentes de cada capa y entre las tres capas, de modo que los procesos que ocurren en una determinada capa embrionaria repercuten en el desarrollo de las demás.
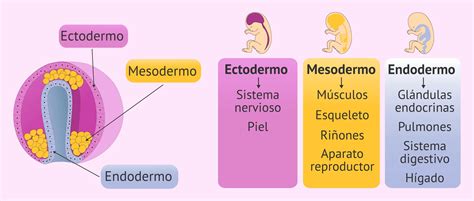
El Ectodermo
El ectodermo es la capa más externa (superficial) del embrión. Por ello, forma parte de las paredes que constituyen el espacio que rodea al embrión: el saco amniótico.
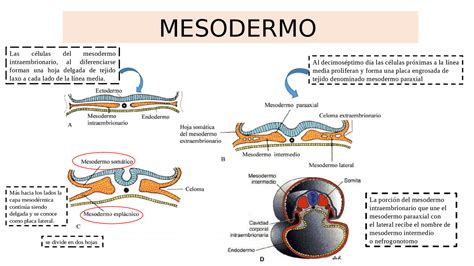
El Mesodermo y su Diferenciación
Aunque todas las estructuras del mesodermo se desarrollan simultáneamente, el componente mesodérmico situado en el eje longitudinal del embrión, la notocorda, es de especial importancia. Originada a partir de la migración rostro-caudal de las células epiblásticas más craneales, la notocorda se extiende a lo largo de todo el embrión, excepto en la zona más rostral donde la placa precordal impide su penetración. La notocorda desempeña un papel fundamental en los procesos de inducción neural sobre la capa ectodérmica suprayacente.
El ectodermo neural es la región ectodérmica longitudinal íntimamente relacionada con la posición longitudinal de la notocorda. Experimentos clásicos han demostrado que la extirpación de la notocorda conduce a la ausencia de diferenciación del ectodermo neural, mientras que la implantación experimental de una segunda notocorda induce la diferenciación de dos regiones ectodérmicas neurales.
Mesodermo Para-axial y Somites
El mesodermo para-axial se caracteriza por su división progresiva en sentido céfalo-caudal, estableciendo la presencia de pares de acúmulos celulares localizados a cada lado de la notocorda, formando así los pares de somites. Un mayor número de pares de somites indica un estadio más avanzado de desarrollo embrionario, siendo un criterio principal para determinar la edad del embrión.
Los somites se diferencian en tres líneas celulares: dermotomo, esclerotomo y miotomo. El dermotomo se relaciona íntimamente con el ectodermo (formador de la epidermis) y dará lugar a la dermis de la piel. El esclerotomo formará estructuras cartilaginosas y óseas, principalmente las costillas y las vértebras del raquis. El miotomo es el componente celular destinado a la formación de las estructuras musculares.
Mesodermo Lateral
El mesodermo lateral se caracteriza por la formación de láminas celulares que se dividen rápidamente en dos capas: una superficial, en relación con el ectodermo, la hoja somatopleura del mesodermo lateral; y otra profunda, en relación con el endodermo, la hoja esplacnopleura del mesodermo lateral. Cada hoja de mesodermo lateral se fusionará con la del lado opuesto a nivel de la línea media del embrión a medida que este se pliega en sentido látero-medial para formar el cuerpo embrionario.
El mesodermo es un claro ejemplo de la especificación del destino durante la gastrulación. Experimentos con trasplantes de segmentos de la línea primitiva entre embriones de pollo y codorniz han demostrado que las células que migran a través de esta estructura adquieren sus compromisos de diferenciación en función del estadio del desarrollo. Por ejemplo, el trasplante de células presomíticas de la línea primitiva a zonas precardíacas resulta en la formación de corazón por parte de estas células y la expresión del gen específico VMHC1.
El Endodermo
El endodermo es la capa embrionaria más profunda, en íntima relación con el saco vitelino. Presenta los cambios morfogenéticos menos llamativos, adoptando una actitud aparentemente pasiva durante el desarrollo inicial, ya que se limita a seguir el proceso de incurvación embrionaria. Da lugar a la constitución del tubo endodérmico, que recorre el embrión longitudinalmente desde la boca primitiva (estomodeo) hasta el ano (membrana cloacal).
Desarrollo del Sistema Nervioso y Circulatorio
Neurulación
Uno de los cambios morfogenéticos más precoces es la diferenciación del ectodermo neural. Alrededor de la tercera semana de gestación se inicia el desarrollo del sistema nervioso, un proceso denominado neurulación. Este incluye la formación de la placa neural y los pliegues neurales, y su cierre para formar el tubo neural, aproximadamente hasta la cuarta semana del desarrollo.
La placa neural se constituye mediante un engrosamiento del ectodermo que se relaciona inicialmente con el nódulo de Hensen y posteriormente con el mesodermo axial (notocorda), prolongándose en sentido cráneo-caudal. Posteriormente, la placa neural se invagina a lo largo de su eje longitudinal para formar el canal neural y, seguidamente, el surco neural, con los pliegues neurales a cada lado. Los pliegues neurales se aproximan y el surco neural se cierra progresivamente, constituyendo el tubo neural.
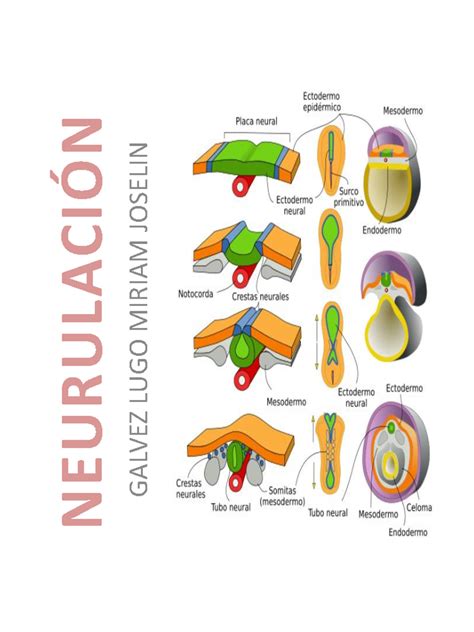
Formación del Encéfalo y Médula Espinal
El tubo neural dará origen al sistema nervioso central: encéfalo y médula espinal. La luz del tubo neural (el conducto neural) formará las cavidades ventriculares y el conducto ependimario, respectivamente. Cuando está finalizando el cierre del neuroporo rostral (el segmento más craneal del tubo neural), tiene lugar el desarrollo del encéfalo. Inicialmente, este sector del tubo determina un proceso morfogenético caracterizado por la formación de las tres vesículas cerebrales primarias: prosencéfalo (cerebro anterior o rostral), mesencéfalo (cerebro medio) y rombencéfalo (cerebro caudal). Estructuralmente, la pared del tubo neural se compone de neuroepitelio cilíndrico pseudoestratificado, grueso, que evoluciona según se configura la estructura histológica.
Desarrollo del Sistema Circulatorio
El sistema circulatorio es la primera unidad funcional en constituirse, siendo el corazón el primer órgano que comienza a funcionar (aproximadamente en la 3ª semana), para suministrar los requerimientos nutricionales y de oxígeno que no pueden ser satisfechos por difusión cuando el embrión se vuelve más complejo. Al comienzo de la fase de gastrulación, las células precardíacas se dirigen hacia la línea media del embrión, se invaginan a través de zonas determinadas de la línea primitiva (situadas justo caudalmente al nódulo de Hensen) y migran rostrolateralmente en distribución bilateral hasta situarse a nivel de las áreas que formarán el corazón.
Desarrollo embrionario en animales
Polaridad Embrionaria y Formación de Ejes
Establecimiento de la Polaridad
Todos los organismos establecen polaridad durante su desarrollo, creando diferencias entre polos opuestos del embrión. Sin embargo, existe controversia respecto al momento de inicio de esta polaridad embrionaria.
En la mayoría de los invertebrados, los primeros síntomas de polaridad y el establecimiento de los ejes embrionarios ocurren antes de la fecundación. Un ejemplo clásico es el de la mosca de la fruta Drosophila melanogaster, donde el RNA mensajero de la proteína bicoid se deposita en un gradiente en el huevo, determinando las futuras regiones anterior (cabeza) y posterior (cola). Este mecanismo, denominado mosaicismo, implica que las células están predestinadas desde el inicio de la segmentación.
En anfibios, los determinantes maternales especifican un eje que se corresponde con el futuro eje dorso-ventral, el eje animal-vegetal. En estos casos, se asume que tanto el óvulo como el embrión en las primeras divisiones no poseen signos de polaridad inicial. Este mecanismo se denomina regulativo, ya que el destino de las células no está fijado y puede modificarse durante el desarrollo.
Polaridad en Mamíferos
La visión aceptada del desarrollo inicial en mamíferos y, por tanto, en la especie humana, es que las células obtenidas durante las primeras etapas son equivalentes y que los primeros signos de polaridad no aparecen hasta la formación del blastocisto o incluso más tarde. Se considera que los ejes embrionarios no se establecen hasta la gastrulación en el epiblasto, mediante interacciones celulares complejas.
Las evidencias que apoyan este modelo regulativo se basan en la plasticidad o potencialidad de las células de los mamíferos antes de la implantación. Sin embargo, se acumulan evidencias experimentales que sugieren una diferencia (polaridad) entre las distintas zonas del embrión en la etapa de blastocisto, mórula e incluso desde la primera división mitótica del cigoto. Si estas evidencias se confirman, implicaría que las blastómeras ya poseen una determinación para producir ciertas regiones del futuro embrión.

Ejes en el Blastocisto y su Origen
El blastocisto de mamíferos presenta un eje claro entre la zona que contiene la masa celular interna (MCI) y la zona opuesta sin MCI. Estas zonas se denominan polo embrionario (Em) y polo abembrionario (Ab), respectivamente, y constituyen el primer eje de polaridad embrionaria distinguible. Este eje divide las células del trofoectodermo en dos grupos: el trofoectodermo polar (cercano a la MCI) y el trofoectodermo mural (alejado de la MCI), con destinos diferentes.
Algunos autores han descrito una asimetría en la disposición de la MCI que la asemeja a un balón de fútbol americano. Se ha establecido una correlación entre esta asimetría y la formación del futuro eje antero-posterior en el embrión. Además, en la MCI, las células más cercanas a la cavidad del blastocele corresponden a las futuras células endodérmicas (ventrales) del embrión, lo que sugiere una correspondencia con el futuro eje dorso-ventral.
Una cuestión importante es si estos ejes observables en el blastocisto se producen de forma aleatoria o si están predispuestos desde etapas previas del desarrollo. La disposición de las blastómeras se explica por una secuencia ordenada de los planos de división celular después de la fecundación, estudiada en embriones de ratón y humanos. Estos planos de división se definen respecto a la posición del corpúsculo polar (CP), estableciendo el eje animal-vegetal (AV) del embrión.
Destino de las Blastómeras
La primera división del cigoto es meridional y produce dos blastómeras simétricas y equivalentes en su contenido de citoplasma animal y vegetal. La segunda división en una de las blastómeras es también meridional, mientras que en la otra es ecuatorial, creando una célula con contenido citoplasmático exclusivamente animal (superior) y otra exclusivamente vegetal. Las células que se sitúan en la periferia darán lugar al trofoectodermo, mientras que las internas formarán la MCI.
Estudios de marcaje celular sugieren que las células están determinadas a formar las zonas del blastocisto más por su origen según las primeras divisiones que por su disposición azarosa en la blástula. Estos análisis, aunque controvertidos debido al intervencionismo experimental, respaldan el modelo de mosaicismo en embriones de mamíferos. La controversia sobre el destino de las blastómeras iniciales ha sido parcialmente zanjada por estudios que muestran que la disposición de las blastómeras condiciona el destino final de las células, y que la relación entre el eje animal-vegetal y el eje embrionario-abembrionario del blastocisto es más sutil de lo que se pensaba.
Polaridad del Cigoto y el Óvulo
Si desde la primera división mitótica las células tienen un destino prefijado, el cigoto debe tener un grado importante de polaridad, posiblemente heredada de una asimetría en el óvulo. Tanto en el óvulo como en el cigoto, la posición del corpúsculo(s) polar(es) denota una polaridad y define el eje animal-vegetal. Este eje coincide con la orientación del primer plano de división y, finalmente, con el destino de las células en el blastocisto.
Aunque se considera que los mamíferos no dependen de componentes citoplasmáticos y que el óvulo y el cigoto son homogéneos, es evidente que deben existir diferencias en la membrana plasmática del óvulo, ya que el espermatozoide no puede penetrar en las cercanías del corpúsculo polar en condiciones fisiológicas. Por lo tanto, aunque no sea observable en el interior del gameto y del cigoto, debe existir una polaridad estructural con respecto al futuro eje animal-vegetal. La localización del corpúsculo polar está determinada por los movimientos que el núcleo realiza en la fase de vesícula germinal, posiblemente relacionados con la maduración ovocitaria y las interacciones entre el ovocito y las células foliculares del ovario.
En resumen, existe una polaridad embrionaria durante las primeras fases del desarrollo de los mamíferos, con algunos signos ya presentes en el óvulo antes de la fecundación. Esta polaridad se confirma y define después de la primera división mitótica, cuando las dos primeras células adquieren destinos diferentes.
tags: #desarrollo #embrionario #patron #dorso #ventral