La capacidad de generar células precursoras de espermatozoides a partir de células madre representa un avance significativo en la comprensión y el posible tratamiento de la infertilidad masculina. Investigaciones recientes han demostrado que es posible inducir la formación de estas células a partir de células madre pluripotentes inducidas (iPSCs), abriendo nuevas vías para la medicina reproductiva.
Investigaciones Pioneras en la Generación de Células Germinales
Avances en Estados Unidos: Células Germinales Primordiales a partir de iPSCs
Investigadores de la Facultad de Medicina de la Universidad Estatal de Montana y la Universidad de Stanford, en Estados Unidos, han logrado que células madre fabricadas a partir de la piel de hombres infértiles adultos produzcan células germinales primordiales. Estas células son las que normalmente se convierten en espermatozoides. Los resultados de esta investigación, publicada en Cell Reports, demostraron que cuando estas iPSCs fueron implantadas en el sistema reproductivo de ratones, se produjo una diferenciación de las células germinales con una eficacia sorprendente.
Los hombres infértiles tratados en este estudio padecían azoospermia, una mutación genética que les impide generar espermatozoides maduros. Este estudio ha permitido una mejor comprensión de las primeras etapas de la reproducción humana.
La Dra. Renee Reijo Pera, exdirectora del Centro de Stanford para la Investigación y la Educación de Células Madre Embrionarias, afirmó: "Vimos una mejor diferenciación de las células germinales en este modelo de trasplante de lo que habíamos visto nunca, estamos sorprendidos por la eficacia."
Éxito en el Reino Unido y Alemania: Espermatozoides Viables que Resultaron en Nacimientos
Un equipo de investigación ubicado en el Reino Unido y Alemania ha tenido éxito transformando células madre en células de esperma viables, dando como resultado un nacimiento. Este avance podría significar el fin del descenso de los índices de infertilidad en los hombres.
El equipo, con base en Newcastle, Göttingen, Mainz y Giessen, consiguió cosechar células madre embrionarias de ratones, transformarlas en esperma viable y fecundar a ratones hembra. Nacieron siete ratones, de los cuales seis sobrevivieron. Tres de estos presentaron anormalidades y murieron posteriormente, pero tres estaban sanos. Los resultados han sido publicados en la revista Developmental Cell.
El equipo reconoció que el esperma procede de una pequeña población de células madre espermatogoniales (SSCs). Según la publicación, se cree que estas células se dividen infinitamente y apoyan la espermatogénesis durante toda la vida en el macho. Utilizando ratones con SSCs agotadas o inactivas, lo que causa infertilidad, el equipo puso a prueba un enfoque diferente, utilizando células madre embrionarias para conseguir el mismo resultado.
Los gametos resultantes fueron inyectados en los óvulos del ratón utilizando una técnica típica de fecundación in vitro humana y después insertados en el útero del ratón.
Investigación en Japón: Espermatozoides Viables a partir de Células Madre Murinas
Un equipo de científicos de la Universidad de Kioto (Japón) también ha logrado producir espermatozoides viables a partir de células madre de ratones. Para ello, extrajeron células madre de embriones de ratones y las diferenciaron en células madre precursoras de los espermatozoides, que luego transplantaron en los testículos de ratones machos infértiles.
La Espermatogénesis: Proceso de Formación de Espermatozoides
La ESPERMATOGÉNESIS: etapas, espermatogonias, diferencias con espermiogénesis
Introducción a la Espermatogénesis
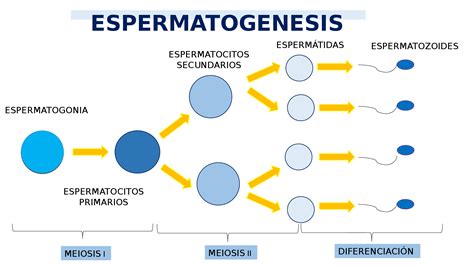
La espermatogénesis es un proceso de mitosis y meiosis por el cual las espermatogonias dan lugar a espermátidas y espermatozoides maduros en los tubos seminíferos del testículo. En un hombre adulto, este proceso se produce continuamente.
Dentro de las espermatogonias, existe un tipo denominado célula madre espermatogonia (spermatogonial stem cell [SSC]) con capacidad de autorrenovarse y de producir células hijas diferenciadas que madurarán hasta espermatozoides. La caracterización de estas células es difícil y no ha sido posible estudiarlas con detalle hasta el desarrollo del trasplante entre especies.
Las SSC son resistentes a una serie de agentes que pueden dañar los testículos, como sustancias químicas o radiaciones en tratamientos oncológicos. En ocasiones, estas células sobreviven cuando otros tipos de células germinales son destruidas, lo que se atribuye a su menor frecuencia de división.
En un corte histológico de un túbulo seminífero de un niño recién nacido se observan túbulos con muy poca luz, junto con células de Sertoli inmaduras y algunas espermatogonias (células madre del testículo), ya que la espermatogénesis no se inicia hasta la pubertad.
Fases de la Espermatogénesis
La espermatogénesis consta de 3 fases principales:
- Fase proliferativa: Las espermatogonias (diploides, con 46 cromosomas) se dividen por mitosis para mantener su "stock" y, a la vez, diferenciarse en espermatocitos primarios.
- Fase meiótica: Durante esta fase, los espermatocitos primarios experimentan la primera división meiótica y forman los espermatocitos secundarios. Estos, a través de la segunda división meiótica, dan lugar a las espermátidas (células haploides y redondas). La fase meiótica dura aproximadamente 24 días en humanos, con 23 días dedicados al estadio de espermatocito primario y solo un día para la segunda división meiótica.
- Espermiogénesis: Las espermátidas se transforman en los espermatozoides finales mediante cambios morfológicos. El citoplasma se desplaza hacia la cola y se elimina junto con los orgánulos y componentes innecesarios (conocido como cuerpo residual o gota citoplasmática). En el ser humano, este proceso dura unos 23 días.
La duración total del proceso de espermatogénesis es de aproximadamente 74 días (desde una espermatogonia hasta el espermatozoide final).
Una vez finalizada la espermiogénesis, ocurre la espermiación, que es la liberación de los espermatozoides desde el epitelio seminífero hacia la luz de los túbulos.
Células de Sertoli
Las células de Sertoli son células somáticas que poseen un citoplasma abundante, ricas en elementos del citoesqueleto y vesículas. Son muy plásticas y contactan con la pared del túbulo seminífero, con células de Sertoli vecinas y con las células germinales.
Además, son importantes recicladoras, ya que se ocupan de eliminar las células que no completan la espermatogénesis y los cuerpos residuales. También facilitan la translocación de las células que inician la meiosis hacia zonas más internas del túbulo, haciéndolas pasar al otro lado de la barrera hematotesticular y migrar hacia la luz del túbulo.
Tipos y Función de las Espermatogonias
Las espermatogonias son células diploides situadas en la base del epitelio seminal, que constituyen el reservorio de células a partir del cual se producen las divisiones celulares que originarán finalmente los espermatozoides. Aunque el modelo más estudiado es el murino, se han producido avances importantes en humanos y bovinos.
En Ratón
En mamíferos no primates, las espermatogonias As (single) son consideradas SSC. Al dividirse, estas células pueden originar:
- Dos nuevas As.
- Espermatogonias Apr, que están predeterminadas a originar el resto de células que conformarán la espermatogénesis.
Cada célula Apr, a su vez, origina dos nuevas células unidas por un puente intercelular, que se mantendrá en su descendencia, para originar las espermatogonias Aal, o espermatogonias alineadas.
Existe un debate sobre si la división de las células madre espermatogonias es simétrica o asimétrica. Experimentos con timidina tritiada sugieren la posibilidad de que una mitosis produzca una célula madre y una espermatogonia Apr, pero no hay pruebas concluyentes.
El gen Zpf145 se ha identificado como clave en el mantenimiento de las SSC en el testículo de ratón, siendo esencial para su mantenimiento y autorrenovación.
La distribución de las espermatogonias en el ratón varía según el estadio:
- En el estadio VIII, las As, Apr y Aal están presentes, aunque las Aal en menor número.
- A partir del estadio X, las Aal proliferan, mientras que As y Apr permanecen más o menos constantes.
- En los estadios II-III, la proliferación se detiene, y las células entran en fase G1-G0.
- En los estadios VII-VIII, casi todas las Aal comienzan una diferenciación al tipo espermatogonia A1, que entra en la fase S del ciclo celular, y alrededor del estadio IX se divide a A2, para posteriormente originar las A3, A4, indiferenciada y B.
El factor neurotrófico derivado de las células de la glía (GDNF), secretado por las células de Sertoli, es un factor clave en la regulación de la diferenciación de espermatogonia As a Apr. A diferencia de la formación de espermatocitos, que tiene mecanismos de regulación exhaustivos, la formación de SSC presenta un escaso control.
El control de la formación de espermatocitos se debe a mecanismos de apoptosis de las A2, A3 y A4, en los que intervienen moléculas de la familia de Bcl-2, como Bax y Bcl-xl.
En Bovino
En bovinos, se ha sugerido una clasificación diferente para las espermatogonias tipo A:
- BSC (basal stem cell): Células redondas con 3 nucleolos irregulares, comparables con las espermatogonias As y Apr del modelo murino.
- ASPC (agregated spermatogonial stem cell): Células de diferentes tamaños.
- CSPC (commited spermatogonial precursor cell): Las espermatogonias más grandes, con un nucleolo central grande, que se corresponden con las espermatogonias A1-A4 diferenciadas del modelo murino.
| Tipo de Espermatogonia | Características en Bovino | Comparación con Modelo Murino |
|---|---|---|
| BSC | Células redondas con 3 nucleolos irregulares | As y Apr |
| ASPC | Células de diferentes tamaños | - |
| CSPC | Células grandes con nucleolo central grande | A1-A4 diferenciadas |
En Humano
Se distinguen tres tipos de espermatogonias:
- Espermatogonias oscuras (Ad): Dentro de las cuales se encuentran las SSC. Se sitúan en íntimo contacto con la membrana basal. Carecen de actividad proliferativa en circunstancias normales, pero entran en mitosis cuando el número de espermatogonias desciende drásticamente. Presentan cromatina muy condensada, una vacuola nuclear pálida, nucleolos pequeños y periféricos, mitocondrias próximas al núcleo, agregaciones de gránulos de glucógeno y cristaloides de Lubarsch.
- Espermatogonias pálidas (Ap): Tienen menor contacto con la membrana basal que las Ad. Presentan un núcleo ovalado y 1 o 2 nucleolos próximos a la membrana nuclear, pero sin vacuola nuclear. Las mitocondrias rara vez forman agrupaciones perinucleares, y las agregaciones de glucógeno son infrecuentes. Sí se observan cristales de Lubarsch. Al dividirse las Ap, se origina la espermatogonia B.
- Espermatogonias B (B): Son las precursoras de los espermatocitos preleptotene. Presentan menor contacto con la membrana basal, gránulos de cromatina muy condensada próximos a la membrana nuclear y un nucleolo situado en la región central. Las mitocondrias están dispersas por el citoplasma.
Fenotipo de las Células Madre Espermatogoniales (SSC)
Fenotipo SSC en el Modelo Murino
Mediante el empleo de técnicas de citometría de flujo e incubaciones con Hoechst 33342 y verapamilo, se han obtenido una serie de características fenotípicas de las SSC en testículo de ratón. El fenotipo antigénico de superficie de SSC sería:
- Positivos: Thy-1+, fracción de integrina α6+, fracción de integrina β1+, CD24+, CD9+, Daz I+, Bax+ y Bcl Xl+.
- Negativos: MHC-I, c-kit, fracción de integrina αv, Sca-1, CD34, CyclinD2 y Bcl2.
Las SSC comparten con otras células madre la ausencia de antígeno de clase I del complejo mayor de histocompatibilidad y la presencia de Thy-1. La expresión de Thy-1 en SSC es inferior a la de las células T y similar a la de las células madre hematopoyéticas. Las células Thy-1+ presentan un fenotipo c-kit, mientras que el c-kit se expresa en las espermatogonias diferenciadas (A1-A4) y es fundamental para su mitosis. La regulación negativa de Thy-1 y la positiva de c-kit podrían ocurrir durante la diferenciación de las SSC.
Una función sugerida para Thy-1 es la inhibición del crecimiento de las SSC, dada su baja tasa de división.
La familia de las integrinas (α6 y β1) juega un papel importante en la migración y anclaje de las SSC a la membrana basal de los tubos seminíferos tras un trasplante.
La proteína CD9, una glucoproteína de membrana de la familia TM4, se expresa en las SSC y está relacionada con la adhesión y migración celular. La proteína Daz I se expresa en las espermatogonias y es fundamental para la diferenciación espermatogénica.
La ciclina D2 parece desempeñar un papel importante en la diferenciación de espermatogonia Aal a A1, mientras que las proteínas apoptóticas Bcl2, Bax y Bcl-xl están implicadas en la regulación de la densidad de las células germinales.
Diferenciación entre SSC y Células SP (Side Population)
Las células SP (side population) son una población de células madre aisladas por primera vez en la médula ósea y luego en otros tejidos. Presentan la particularidad de emitir fluorescencia con tinciones y diferenciarse en células de diferentes tejidos. Aunque se identificó una población de células SP en el testículo de ratón, estudios con citometría de flujo, tinción de Hoescht 33342 y verapamilo permitieron definir un fenotipo distinto para esta población (Sca-1+, Thy-1, c-kit, CD34+/, CD43, CD45+/, MHC-I+). Se concluyó que las SSC y las SP son dos poblaciones celulares diferentes, y que las SP testiculares no contienen SSC, ya que no mostraron actividad espermatogénica tras el trasplante.
Fenotipo SSC en el Modelo Bovino
La investigación en el modelo bovino está menos avanzada que en el murino, pero se han logrado caracterizar fenotípicamente las células madre espermatogonias. Las espermatogonias tipo A se caracterizan por reaccionar con la lectina DBA. Dentro de las espermatogonias DBA+, se aislaron dos poblaciones celulares: una más grande con una fuerte reacción positiva a c-kit (indicando diferenciación) y otra más pequeña sin esta reacción (conteniendo células madre espermatogonias).
Otra característica fenotípica de las SSC en bovinos es la influencia del GDNF, ya que la adición de GDNF exógeno a un cocultivo de células aumenta el número de SSC.
Aplicaciones Clínicas y Retos Futuros
Células Madre con Pluripotencia Inducida (iPSCs)
Las células de nuestro cuerpo pueden retornar al estado de célula indiferenciada, capaz de derivarse a cualquier tipo celular. Este hallazgo es de gran importancia, pero el reto radica en proporcionar a estas células las señales adecuadas para dirigir su especialización hacia las células del tejido de interés.
La importancia de los estudios recientes reside en que, una vez desdiferenciadas las células humanas, se inyectaron en el testículo de ratón, donde se produjo espontáneamente la división celular y las transformaciones necesarias para dar lugar a un espermatozoide inmaduro. Los investigadores explican que no se alcanzó el estado final de espermatozoides funcionales debido a las diferencias moleculares entre la formación de espermatozoides murinos (de ratón) y humanos.
La investigación también incluyó la recolección de muestras de tejido de hombres azoospérmicos, cuyas células desdiferenciadas inyectadas en los túbulos seminíferos de ratones también provocaron la diferenciación a espermatozoides inmaduros.
Importancia del Estudio de Espermatogénesis Inducida
La autora principal del estudio, Renee Reijo Pera, defiende que sus "resultados son los primeros en ofrecer un modelo experimental para estudiar cómo se desarrollan los espermatozoides". Este modelo in vivo de producción espermática a partir de células desdiferenciadas permitirá profundizar en el tratamiento de los problemas de infertilidad masculina relacionados con defectos en la producción de espermatozoides.
En el futuro, podría ser posible trasplantar células madre de pluripotencia inducida en túbulos seminíferos de varones infértiles, especialmente aquellos con defectos genéticos que resultan en la pérdida de un fragmento cromosómico del cromosoma Y, no congénito, sino posterior a las primeras divisiones embrionarias.
El profesor Karim Nayernia, investigador jefe de uno de los equipos, destacó en una entrevista con el periódico Independent que la investigación es particularmente importante para comprender mejor el proceso biológico de producción de esperma. La investigación tiene más aplicaciones que una simple fecundación in vitro; podría proporcionar un tratamiento, considerando que más de la mitad de los tratamientos de infertilidad se dirigen a la infertilidad masculina.
"Si sabemos más acerca de cómo las células madre espermatogoniales se convierten en células de esperma, esto podría traducirse en tratamientos para hombres cuyo esperma es disfuncional," dijo el profesor Nayernia. Por ejemplo, se podrían aislar las células espermatogoniales de un paciente mediante una biopsia testicular simple, estimularlas en el laboratorio para que se conviertan en esperma funcional y volver a trasplantarlas al paciente.
Consideraciones Éticas, de Seguridad y Legales
El desarrollo de estas técnicas ha generado alarma en algunos grupos preocupados por la inviolabilidad del embrión. Sin tener en cuenta las implicaciones éticas, los aspectos de seguridad y legales de esta investigación podrían tardar años en resolverse. No obstante, la técnica podría, en teoría, poner fin a la especulación sobre cómo reducir la caída de los índices de esperma. Se estima que el 30 por ciento de los hombres son ahora subfértiles con problemas de fertilidad, mientras que otro dos por ciento son completamente infértiles.
Actualmente, en humanos, no se ha logrado la diferenciación de este tipo de células madre inducidas en gametos idénticos a los generados in vivo, sino solo similares. Incluso, existe especulación sobre la posibilidad de modificar genéticamente óvulos generados artificialmente para prevenir o eliminar enfermedades en la descendencia mediante técnicas de edición génica como CRISPR. Sin embargo, aunque la investigación avanza, la aplicación clínica en los laboratorios de reproducción asistida aún está lejos, requiriendo grandes avances en eficacia, seguridad y reproducibilidad.
Renee Reijo también explicó que los resultados y enfoques podrían proporcionar nuevas estrategias en el diagnóstico y asistencia de aquellos que sufren infertilidad genética o secundaria, al igual que podrían ser utilizados para tratar enfermedades degenerativas o relacionadas con la edad. "Nuestro estudio sugiere que el uso de células madre puede servir como material de partida para el diagnóstico de cáncer de células germinales y generando potencialmente células germinales."
Estudio de la Meiosis en el Varón
Análisis Citogenético
El estudio de la meiosis en el hombre adulto se puede realizar a nivel citogenético a partir del análisis del tejido testicular obtenido mediante biopsia. En el análisis citogenético, se obtienen y analizan las células precursoras de los espermatozoides en las diferentes fases de la meiosis, destacando la profase I, la metafase I y la metafase II.
- En la profase I se observan los 46 cromosomas semidescondensados, con el apareamiento de los cromosomas homólogos iniciado (leptoteno-zigoteno) y la condensación de los filamentos haciéndose más evidente.
- En la metafase I se diferencian los 23 pares de cromosomas más condensados, en forma de bivalentes unidos por los quiasmas. Esta fase permite detectar anomalías de recombinación.
- La metafase II es una fase muy corta, por lo que suele ser difícil de encontrar y diagnosticar.
Este tipo de estudio permite conocer si el reparto de los cromosomas se desarrolla correctamente a lo largo de la meiosis o de forma incorrecta. Un reparto erróneo puede resultar en espermatozoides con un número anormal de cromosomas: diploides (doble dotación cromosómica) o aneuploides (un cromosoma de más o de menos).
Análisis Histológico
El estudio también puede realizarse desde el punto de vista histológico, analizando el tejido testicular. Es crucial que los cortes sean completamente transversales y la tinción correcta para diferenciar morfológicamente cada una de las células del túbulo.
Patologías Asociadas con la Meiosis y Consecuencias
En un epitelio sano, se observan unas 4 o 5 capas de células germinales. En la base del túbulo se distinguen los núcleos pequeños y heteropignóticos de las espermatogonias, y a medida que avanza la meiosis, las células se encuentran más cercanas a la luz del túbulo.
Si se observan pocas capas de epitelio seminífero, puede ser un síntoma de una espermatogénesis ineficiente. En algunos casos, se observan grupos de células que se descaman del túbulo seminífero, perdiendo su capacidad de proliferación. Un tejido testicular mal estructurado se relaciona con infertilidad (oligozoospermia).
Otra patología es el síndrome de Sertoli-only, caracterizado por la presencia exclusiva de células de Sertoli que colonizan la totalidad del túbulo.
Se ha observado que los pacientes con meiosis alterada suelen presentar también alteraciones en el seminograma.
tags: #celulas #precursoras #de #espermatozoides