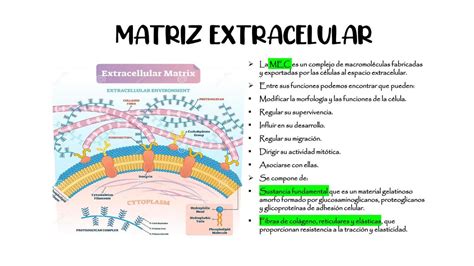
La migración celular es posiblemente la función más conocida que se le asigna a la matriz extracelular (MEC) durante la embriogénesis. Sin embargo, poco se sabe de cómo la MEC regula la migración celular in vivo, ya que su estudio en el ambiente tridimensional de un organismo en desarrollo es complicado. Por ello, la mayoría del conocimiento que se tiene sobre el papel de la MEC en embriogénesis deriva de experimentos realizados con células u órganos en cultivo.
La Matriz Extracelular en la Migración Celular Embrionaria
Fuentes de MEC y Regulación de la Migración Celular
Esta función de la MEC en la migración celular durante la embriogénesis ha sido clásicamente atribuida a la MEC expuesta en el sustrato, conocida como matriz exógena. No obstante, las células también pueden usar una MEC endógena, secretada de manera autocrina. Sin embargo, se desconoce si esta capacidad de las células de usar su propia matriz para migrar se utiliza durante la embriogénesis.
Estudio en Drosophila melanogaster: Un Modelo para la Migración de Macrófagos
Por ello, en una tesis doctoral se han usado los macrófagos de Drosophila melanogaster como sistema modelo para el estudio in vivo del papel de la MEC en la migración celular durante la embriogénesis. En concreto, esta tesis se ha centrado en los siguientes aspectos:
- Se ha estudiado el papel de las lamininas, principales componentes de la MEC, en la migración de los macrófagos in vivo.
- Se ha analizado la contribución de las lamininas endógenas y exógenas en este proceso de migración.
- Se han examinado los mecanismos moleculares y celulares por los cuales las lamininas regulan la migración de los macrófagos.
Hallazgos Clave sobre las Lamininas y Otros Componentes
Los resultados obtenidos demuestran que los macrófagos usan lamininas endógenas y exógenas para su migración a través del embrión. Si bien, se propone que estas dos fuentes de lamininas ejercen funciones distintas: la laminina exógena tendría un papel más estructural, implicado en la correcta formación del espacio intersticial por el cual migran los macrófagos. También se muestra que las lamininas regulan la direccionalidad y la estabilidad de las proyecciones de macrófagos al migrar. Además, se implica a las Rho GTPasas Rabs en la secreción de las lamininas. Por último, se ha analizado el posible papel de otros componentes de la MEC, como Nidogen y Colágeno IV, en la migración de los macrófagos.
Desarrollo celular embrionario
La Matriz Extracelular en las Membranas Fetales Humanas
Membranas Fetales: Tejidos Extraembrionarios Esenciales
Todos los vertebrados, desde los ovíparos hasta los vivíparos, tienen tejidos extraembriónicos accesorios los cuales contienen al feto, conocidos como membranas fetales. Estos tejidos comprenden tres láminas germinales primarias, que nunca son inervadas, y existen sólo como tejidos embriónicos accesorios. Ellas son genéticamente idénticas al feto, pero tienen un tiempo de vida limitado, estando presentes sólo hasta el punto en el que el feto se desarrolla lo suficiente para convertirse en un individuo funcional. La placenta puede ser considerada como una región especializada de las membranas; se puede pensar que las membranas son apéndices de la placenta. Ambos son puntos altamente especializados de la interacción materno-fetal, las cuales parecen tener su mayor importancia, tanto en el mantenimiento del embarazo como en el parto, en los vertebrados superiores.
Importancia Biomecánica y Desafíos en el Embarazo Humano
El objetivo de la investigación será las membranas fetales extraplacentarias, amnios, corion y la decidua materna contigua, las cuales también han sido descritas como una membrana por sí mismas. La marcada diferencia entre los tipos morfológicos de las membranas presentes en todos los vertebrados, puntualiza la necesidad de enfocarse en una sola especie. El embarazo y el parto en los humanos representan un grupo único de problemas particularmente para las membranas fetales las cuales forman un continente biomecánico ajustable al crecimiento y movimientos del feto dentro de un bípedo erguido. Quizá la evolución de esta postura ha llevado a una dificultad relativa para el nacimiento en nuestra especie.
La postura erguida es significativa para las membranas fetales y representa para ellas un mayor reto mecánico que en otras especies. Durante el embarazo existe la necesidad que las membranas fetales sean lo suficientemente fuertes y elásticas para soportar un estiramiento del doble de su tamaño al momento del término del embarazo y simultáneamente resistir los movimientos fetales vigorosos. Por otro lado, la construcción de una estructura gruesa pero elástica, como son los tejidos elásticos normales, pudieran prevenir la necesidad de una ruptura programada de las membranas, lo cual es un evento normal durante la primera etapa del trabajo de parto.
Para la mayoría de los embarazos, el trabajo de parto comienza a las 38-42 semanas de gestación en presencia de membranas intactas. La ruptura a término antes de la aparición de contracciones uterinas regulares ocurre en cerca de 2 % - 18 % de los embarazos. Esto puede ser una imprecisión fisiológica de la sincronización con algunas consecuencias adversas. En marcado contraste, la ruptura prematura en embarazos pretérminos precede en un 30 % - 40 % de todos los partos y conlleva riesgos significativos para el producto. Es un evento patológico raro y no existe en otros vertebrados. Además, la comprensión de los componentes estructurales básicos de las membranas fetales humanas, y cómo ellos se adaptan a las necesidades de cambio cuando el contenido uterino aumenta, es fundamental para el eventual control de este gran problema de salud. También nos hace preguntar sobre lo apropiado de los modelos animales para los estudios del embarazo y el parto en los humanos. Las membranas fetales humanas son tejidos relativamente accesibles, durante ciertos momentos del embarazo. Usar muestras de tejidos pretérminos los cuales son patológicos puede representar una desventaja, pero esto es opacado por la relevancia clínica de los datos obtenidos. Sin embargo, esta revisión se enfoca tanto como es posible en el conocimiento actual de los principales componentes de la matriz extracelular de las membranas fetales en embarazos a término en humanos.
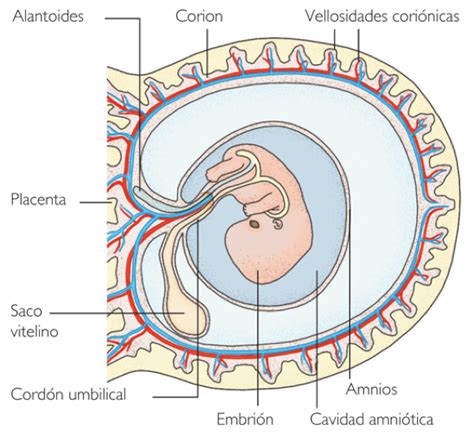
Organización y Ultraestructura de las Membranas Fetales
Están disponibles muchas descripciones de la formación de la placenta y las membranas fetales. Sin embargo, la complejidad de la formación enmascara los orígenes del amnios, del corion y su desarrollo. El corion se forma del blastocisto implantado en el polo que apunta hacia la cavidad endometrial, la cual es cubierta por el corion frondoso y la decidua capsular. El flujo de sangre se restringe en esta área y las vellosidades degeneran formando el corion avascular. El amnios se forma separadamente del componente epitelial y mesenquimal y se fusiona con el mesodermo del corion formando el "corioamnios". Cuando el embrión crece, el remanente de la decidua vascular adherida al corion se opone a la decidua parietal materna. Además, el útero ofrece soporte a las membranas fetales las cuales se alinean completamente con éste a la mitad del embarazo.
Capas del Corioamnios
La organización normal y la anatomía microscópica del corion y amnios humano han sido descritas en detalle por los estudios clásicos de Bourne y es difícil mejorarlo. El diagrama de Bourne muestra la organización general de este sistema de multicapas:
- La capa más interna es el epitelio amniótico, en contacto directo con el líquido amniótico.
- Éste se superpone a la membrana basal la cual se opone a la capa compacta, que varía en espesor.
- Sobre ésta se ubica la capa de fibroblastos, una región de células mesenquimales dispersas.
- La capa esponjosa es rica en proteoglicanos los cuales permiten el paso del agua, facilitando que el amnios pueda deslizarse sobre el corion, un mecanismo que ha sido propuesto como un sistema de reparación mecánica a corto plazo.
La matriz extracelular coriónica subyacente y las células del citotrofoblasto en el embarazo a término están firmemente adheridas a la decidua materna y cuando se usan marcadores específicos para las células deciduales es común encontrar a estas células infiltrando a través del citotrofoblasto hacia la membrana seudobasal.
Función de la MEC en la Resistencia Mecánica
La ultraestructura de las membranas amnióticas ha sido objeto de un gran número de estudios a nivel de la microscopia electrónica de transmisión. Los principales componentes son células y matriz extracelular. Las primeras son responsables de la síntesis, degradación y recambio de la última. La matriz extracelular en cambio influye sobre las funciones de los componentes celulares durante el embarazo. La mayor fuerza tensil del amnios es dada por los colágenos de la capa compacta más allá del epitelio amniótico. Sin embargo, este arreglo de los colágenos más allá del intersticio y conectados a los colágenos de la membrana amniótica basal da fuerza extra a los tejidos expuestos a las fuerzas mecánicas tanto de tipo repetitivo, como en los vasos sanguíneos, o de tipo continuo como el que es necesario cuando las membranas son estiradas en el embarazo a término.
Principales Componentes Extracelulares: Colágenos
Clasificación y Ensamblaje de los Colágenos
Se conocen hasta el momento por lo menos 19 tipos genéticamente distintos de colágenos, los cuales son codificados por lo menos por 30 genes formando una familia especializada de glicoproteínas estructurales. La clasificación como colágeno está basada sobre si contiene una secuencia repetida de Gli-X-Y y si éste forma un componente integral de la matriz extracelular. El conocimiento de cómo se ensambla la triple hélice de colágeno y cómo interactúa con otros componentes extracelulares de la matriz se ha incrementado rápidamente en años recientes y ha sido discutido con detenimiento.
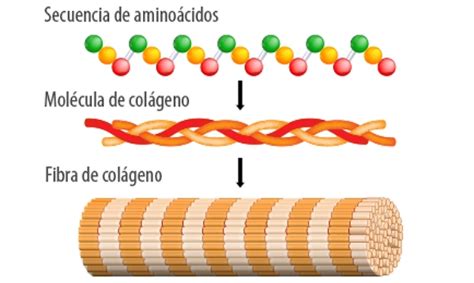
Colágenos Intersticiales y de Membrana Basal
Los colágenos son los principales componentes estructurales de las membranas fetales como se ha demostrado tanto por estudios de extracción y caracterización como su inmunolocalización específica dentro de estos tejidos. La mayor fuerza tensil de las membranas fetales es provista por los colágenos intersticiales, tipo I y III, junto con pequeñas cantidades de los tipos V, VI y VII en la capa compacta sobre la membrana amniótica basal. Esta última es una membrana basal clásica del colágeno tipo IV, la cual da una base para el ensamblaje de otros componentes de la membrana basal: laminina, entactina/nidógeno y el proteoglicano heparan sulfato.
Colágenos Menores y de Anclaje
Sin embargo, los componentes colágenos cuantitativamente menores, tipos V, VI y VII, en la región compacta son importantes para el reforzamiento debido a que los tipos V y VI forman fibrillas heterotípicas con los colágenos tipo I y III. El colágeno tipo V es considerado un colágeno fibrilar menor, pero su distribución en la vecindad inmediata de la membrana amniótica basal sugiere que también realiza funciones de anclaje en este punto. Aunque el colágeno tipo IV está asociado con las membranas basales, pero en este caso es parte integral de la lámina basal del epitelio amniótico y de la membrana seudobasal del corion, Malak y col. demostraron su amplia distribución a través de estos tejidos. Se ha sugerido que el colágeno tipo IV puede también tener un papel en el desarrollo y mantenimiento de la estructura de la matriz. El colágeno tipo IV también se encontró distribuido a lo largo de todos estos tejidos apoyando la visión que este colágeno está probablemente diseminado en los tejidos conectivos.
El colágeno tipo VII da fijación adicional, uniendo la lámina basal del epitelio amniótico a la matriz extracelular subyacente en forma de fibrillas de anclaje. Este es un colágeno particularmente largo el cual forma dímeros antiparalelos, el principal constituyente de las fibrillas de anclaje. El colágeno tipo XIV, un colágeno asociado a las fibrillas, también está presente en la matriz del amnios-corion y la membrana basal de la decidua. Estos colágenos menores pueden ser importantes para la estabilidad mecánica de los tejidos y dan elasticidad frente al estiramiento que sufren durante la parte final del embarazo. No debe olvidarse que las estructuras intracelulares también dan fuerza potencial y capacidad de reparación al amnios-corion, esto ha sido descrito con detenimiento y está más allá del objetivo de este estudio.
Síntesis de la MEC y Función Celular en las Membranas Fetales
Contribución de las Células Epiteliales y Mesenquimales
Los primeros informes describían que el colágeno tipo I y III estaban asociados con las células del citotrofoblasto. Estudios más recientes, sin embargo, no han demostrado que éste sea el caso. El colágeno tipo IV, por otra parte, ha sido identificado en asociación con estas células en todos los estudios de inmunolocalización. Las células epiteliales amnióticas sintetizan tanto el colágeno constituyente de su lámina basal (tipo IV) como los colágenos del estroma intersticial (tipo I y III) de la lámina compacta. Se ha demostrado que la producción de estos componentes de la matriz extracelular continúa hasta el término del embarazo.
Estudios recientes han usado preparaciones similares de células epiteliales amnióticas, además de cultivos de células mesenquimales confluyentes que se aislaron del estroma y se usaron en paralelo. Se demostró que la síntesis de los colágenos intersticiales era una función primaria, pero no exclusiva, de las células mesenquimales más que de las células epiteliales. Se observó que la mayor actividad de síntesis del mesénquima para el colágeno intersticial era temprana en el embarazo, declinando al término de éste. Sin embargo, esto va en paralelo con una disminución de la densidad de las células mesenquimales por unidad de área de amnios. El cultivo de tejidos a término, por otra parte, los cuales mantienen la matriz y las células intactas y en condiciones similares a la situación in vivo, sugiere que tanto las células epiteliales como mesenquimatosas contribuyen a la producción, por lo menos, del colágeno tipo III.
La función de las células aisladas in vitro tiene que probar ser iguales a aquellas in vivo antes que se puedan obtener conclusiones firmes. Además, se han reportado cambios en el fenotipo de las células epiteliales amnióticas cuando se cultivan con diferentes condiciones y la producción in vitro de factores de crecimiento, citokinas, prostaglandinas u hormonas por las células de la decidua depende mucho de las condiciones usadas. Sin embargo, parece que aun en embarazos a término, la capacidad de reforzar la capa compacta por síntesis de colágenos intersticiales tanto por la producción epitelial como por la producción mesenquimal está presente.
El Corion: Funciones de Protección y Regulación Hormonal
El corion tiene una función tanto protectora contra el rechazo inmunológico como también es responsable de la desactivación de un número importante de hormonas, prostaglandinas, ocitocina y endotelina-1 producidas localmente. La matriz extracelular asociada con las células citotrofoblásticas coriónicas pueden ser importantes para la fijación de la decidua hasta el parto, separación y desprendimiento que ocurren en la estría decidual, dejando a la lámina decidual capsular / parietal adherida al corion. La existencia de cavidades irregulares en la placa basal de la necrosis o apoptosis de células trofoblásticas o deciduales incluidas en la matriz de fibrinoide, es un factor que debilita la región a ser desprendida, las cuales envejecen y mueren en la unión feto-materna. Es posible que los componentes de anclaje envejecidos o muertos, contribuyan a originar estas cavidades. Este tejido ha sido preparado para un evento similar al que ocurre al final del ciclo menstrual.
La Decidua y su Matriz Extracelular
Composición y Punto de Contacto Materno-Fetal
La decidua humana es una mezcla de células maternas de diferentes orígenes junto con abundante matriz extracelular. El punto de contacto entre el citotrofoblasto y la decidua materna es una región importante, debido a que los nutrientes que necesita la decidua difunden a través del corion avascular, al mismo tiempo que se necesita mantener una barrera inmunológica. Las verdaderas células deciduales derivan del estroma uterino materno, pero la comunicación intercelular está ausente en los humanos y tiene uniones inusuales donde las proyecciones de las células se prolongan y se unen a las mismas células deciduales.
Anclaje y Señalización Paracrina
Hay una gran población (más de 47 %) de células derivadas de la médula ósea en la decidua humana en el embarazo a término. Estas células menores tienen conexiones especializadas a las células deciduales, sugiriendo una comunicación cruzada entre diferentes tipos de células. Alrededor de cada célula decidual con sus macrófagos y linfocitos que las acompañan tienen una membrana basal pericelular distinta. La matriz en esta posición puede ayudar al anclaje de los macrófagos migratorios a la decidua. También puede representar un papel significativo en la dirección del flujo de las señales paracrinas desde y hacia las células deciduales; por ejemplo, la prolactina decidual es transportada a través de las membranas fetales hasta el líquido amniótico.
tags: #matriz #extracelular #en #el #desarrollo #embrionario