Definición y Proceso General
La embriogénesis vegetal es el conjunto de procesos fisiológicos que conducen a la transformación de una sola célula, el cigoto, en un individuo multicelular más complejo, el embrión, contenido en la semilla madura. Este proceso requiere una fina regulación de multitud de elementos de desarrollo que conducen a la elaboración de morfologías básicas (morfogénesis), el establecimiento de estructuras funcionalmente organizadas (organogénesis) y la diferenciación tisular.
Durante la embriogénesis, la planta tiene que superar varias etapas. Una de ellas es establecer el plan corporal básico del esporófito, el cual es reestructurado cuando se rompe la dormancia. Para establecer este plan, inicialmente se dan divisiones radiales que forman tres sistemas tisulares básicos: el dermal, el basal y el vascular. Luego se procede a las divisiones axiales que forman el eje apical (tallo)-basal (raíz).
La embriogénesis en las plantas se refiere al proceso de desarrollo mediante el cual se forma y desarrolla el embrión dentro de la semilla. Es la fase temprana en el ciclo de vida de una planta, comenzando desde un cigoto unicelular, que se forma cuando el gameto masculino (espermatozoide) fertiliza al gameto femenino (óvulo), y progresa hasta la formación de una estructura multicelular que eventualmente se convierte en el embrión maduro. Durante la embriogénesis, el cigoto experimenta una serie de divisiones y diferenciaciones que conducen a la formación de varios tejidos y órganos de la planta.
Establecimiento de la Polaridad y Plan Corporal
En algas pardas, que son un sistema modelo utilizado por su fertilización externa y la ausencia de múltiples capas de tejido en comparación con las angiospermas, la primera división asimétrica del embrión establece la polaridad. Esta división produce una célula más pequeña que forma el rizóide, homólogo a la raíz y ancla el resto de la planta, y otra célula más grande que genera el tallo, el cuerpo principal del esporófito.
El punto de entrada del esperma establece la polaridad de la planta, fijando dónde será el rizoide terminal del eje apical-basal, perpendicular al plano de la primera división. La F-Auxina se acumula en el rizoide terminal. Sin embargo, la fijación de la polaridad puede ser anulada por la gravedad o la luz, estímulos que establecerían una nueva posición para la división celular.
Después del establecimiento de la polaridad, vesículas secretoras se mueven hacia el rizoide terminal del cigoto, conteniendo material para el desarrollo del rizoide y la formación de una pared celular con una composición diferente de macromoléculas.
En angiospermas, el establecimiento del plan corporal también comienza con una división asimétrica. La primera división da lugar a una célula basal y una terminal. La célula terminal da lugar al embrión, y la célula basal, formada más cerca del micrópilo, da origen al suspensor, que sirve como conducto de nutrientes.
El proceso que lleva desde el zigoto hasta el embrión se denomina embriogénesis, durante la cual se establecen dos ejes: el apical-basal (tallo-raíz) y el radial (interno-superficial). La primera división del zigoto produce dos células, una apical y otra basal. La apical dará lugar al embrión. La basal es grande y vacuolada, y dará lugar al suspensor, una estructura encargada de anclar el embrión a la zona del micropilo y que llevará nutrientes al embrión. Los derivados de la célula apical darán primero a una masa de células redondeada denominada proembrión, que se alargará y crecerá en tamaño. Tras ello, comenzará la diferenciación celular. Se forma el protodermo en la periferia, el meristemo fundamental y el procambium internamente. Estos son los tres meristemos primarios.
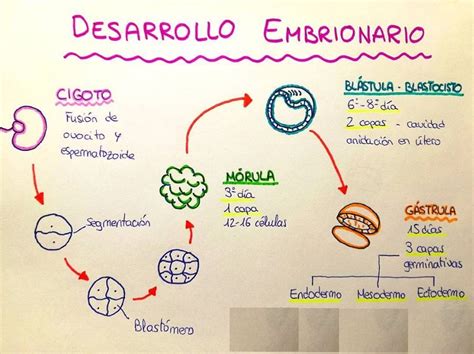
Estructuras del Embrión y Meristemos
El meristemo apical del tallo y el meristema apical de la raíz son cúmulos de células madre que continúan en la planta postembrionaria y dan lugar a la mayoría del cuerpo del esporófito. El meristemo de la raíz se origina parcialmente de la hipófisis en algunas especies. El resto de las estructuras del esporófito se originan del embrión.
El embrión maduro, antes de que la semilla entre en dormancia, consta de varias partes: un epicótilo (encima de los cotiledones) que tiene un meristemo apical, los dos cotiledones, y el hipocótilo (debajo de los cotiledones) que tiene los primordios del tallo y de la raíz. Si se observa una zona de tallo más diferenciada en el epicótilo se denomina plúmula, y si se observa de raíz en el hipocótilo se llama radícula.
Cotiledones y Diversidad Embrionaria
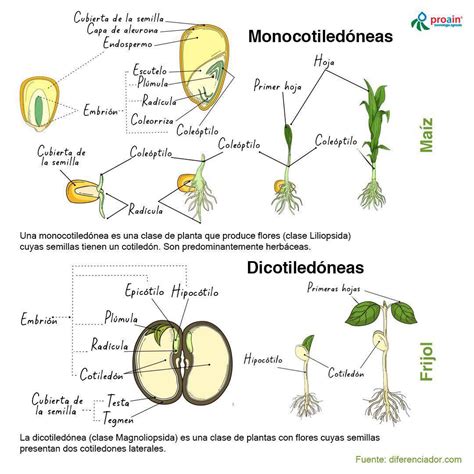
La mayoría de las angiospermas son dicotiledóneas o monocotiledóneas. En el caso de las dicotiledóneas, el zigoto se divide en dos por medio de un tabique longitudinal, separando los futuros cotiledones. Los cotiledones son estructuras a modo de hoja que sirven de almacén o son absortivas y están conectados vascularmente con el embrión. Pueden almacenar sustancias de reserva para la germinación y entonces suelen tener un aspecto carnoso.
En monocotiledóneas, y algunas dicotiledóneas, el cotiledón es absortivo y las reservas de alimento del embrión se almacenan en otra parte de la semilla, de manera que el cotiledón es un estado intermedio en la transferencia de sustancias de reserva desde la semilla al embrión. Si los cotiledones tienen tejido de reserva no contienen meristemo, pero si van a realizar la fotosíntesis mantienen algún potencial meristemático. Los cotiledones están unidos al eje embrionario en un punto llamado nodo y se abren hacia afuera como un libro.
El desarrollo de los embriones de monocotiledóneas es similar al de dicotiledóneas, pero con el embrión más alargado, debido sobre todo a la presencia de un solo cotiledón. Las gramíneas tienen un embrión con un epicótilo que tiene un meristemo apical rodeado por primordios foliares que forman una vaina llamada coleóptilo, y una raíz con una vaina llamada coleorriza.
Sustento Molecular y Genético
Si bien la morfología del embrión vegetal es notablemente simple, el sustento molecular que lo permite es complejo. La expresión genética en embriones es amplia: se ha estimado que los RNAs embrionarios de *Nicotiana tabacum* (la planta de tabaco) son unos 20.000 genes, un número equivalente al expresado en tejidos diferenciados como hojas o anteras. No obstante, se ha estudiado qué genes se expresan en mayor medida durante la embriogénesis, y corresponden a los codificantes para proteínas de almacenamiento de la semilla.
En un reciente congreso en Barcelona se revisaron los datos sobre los genes y los mecanismos moleculares que gobiernan el proceso de embriogénesis. Ya comienzan a identificarse estos genes y las señales que utilizan las células del embrión para comunicarse durante el proceso en el que deben comenzar a definirse los distintos órganos. En el embrión final estarán definidos la raíz, el tallo y las hojas que se formarán cuando la planta germine. Ahí, por tanto, se decide la organización del conjunto del organismo.
Endospermo y Cubiertas de la Semilla
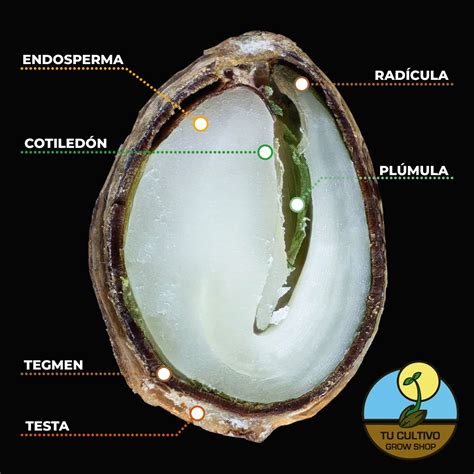
El endospermo es un tejido nutritivo que se encuentra a un lado del embrión o rodeándolo. En el caso de las angiospermas, procede de la fusión de un núcleo generativo con los dos núcleos centrales del saco embrionario, formando un tejido triploide llamado endospermo secundario. En el caso de las gimnospermas, este tejido nutritivo es haploide y se denomina endospermo primario.
El endospermo es un tejido de reserva que proporciona nutrientes al embrión y a las primeras fases del desarrollo de la planta. Las células nutricias almacenan granos de almidón o proteínas que pueden formar gránulos amorfos llamados glútenes o complejos proteicos cristalizados llamados granos de aleurona. Las semillas que contienen endospermo en estadios maduros se denominan semillas endospérmicas o albuminosas, mientras que hay algunas que lo consumen en los primeros estadios de la maduración y se denominan semillas no endospérmicas o exalbuminosas.
Las envueltas de la semilla son de origen materno y surgen a partir de los tejidos que rodean al óvulo. La formación de la cubierta está inhibida antes de la fecundación, y la fecundación elimina esta inhibición, permitiendo el desarrollo de la cubierta. La cubierta se origina principalmente a partir de los tegumentos interno y externo del rudimento seminal, los cuales se convertirán en el tegmen y la testa de la semilla, respectivamente. Normalmente, tegmen y testa están unidos y es difícil separarlos, excepto en algunas plantas como las judías. Conjuntamente se denominan epispermo o cubierta seminal.
El tegmen es normalmente delgado y flexible, mientras que la testa es dura. En la superficie de la testa se sitúa una capa de células a modo de epidermis que desarrollan una cutícula que supone una barrera física para el agua y agentes externos, pero es semipermeable a los gases. A veces, en la cubierta, se pueden encontrar moléculas denominadas genéricamente defensinas, que son repelentes o tóxicas frente a patógenos o herbívoros. En algunas especies hay un elemento adicional de protección consistente en sustancias tóxicas.
En general, las células de las envueltas se convierten en células con paredes celulares más gruesas, y algunas se diferencian en esclereidas de diversos tipos, aunque también puede haber capas de aerénquima y clorénquima entremezclados. Histológicamente hay una gran variedad en la organización de la cubierta de las semillas según las diferentes especies de plantas, y estas envueltas protectoras pueden proceder de las células de la nucela o incluso del saco embrionario.
En Arabidopsis, la cubierta de la semilla consta de 5 capas: tres internas y dos externas que forman el denominado tegumento. El endotelio es la más interna, las otras dos siguientes se funden a medida que se desarrolla la semilla. Las dos externas son las que sufren una mayor diferenciación y forman una capa subepidérmica y otra epidérmica. La subepidérmica forma una pared gruesa, mientras que la epidérmica forma una capa de mucílago. En las leguminosas, la cubierta está formada por muchas capas que incluyen macro y osteoesclereidas en las capas externas y parénquima en las internas.
Las cubiertas protectoras adquieren una gran consistencia la mayoría de las veces, fundamentalmente por la acumulación de esclereidas, aunque otras veces pueden llegar a ser carnosas. Las testas, tanto de monocotiledóneas como de dicotiledóneas, están marcadas normalmente con relieves y texturas, a veces estructuras a modo de alas. Una característica de la cubierta es que debe mantener protegida a la semilla del exterior, pero al mismo tiempo debe sentir el ambiente para permitir la germinación cuando las condiciones son adecuadas. En muchas especies, la capa epidérmica produce una cutícula que hace de barrera impermeable entre el exterior y la semilla, que no pueden cruzar ni virus ni bacterias, pero sí débilmente los gases.
En la superficie de las semillas siempre hay una cicatriz denominada hilio, que corresponde al punto de unión del rudimento seminal con el funículo, pequeño cordón que une la semilla a la pared del ovario. También hay una pequeña abertura denominada micropilo, punto de entrada del tubo polínico, por donde puede entrar el agua para favorecer la germinación.
Funciones y Adaptaciones de la Semilla
Hay semillas, como las de las orquídeas, que pueden ser de unas 200 µm de diámetro. Estas semillas no tienen sistema vascular y los nutrientes y señales hormonales viajan por vía apoplástica. En semillas medianas y grandes se desarrollan sistemas vasculares, más complejos cuanto más grande es la semilla. En algunas semillas los haces vasculares terminan en la unión del funículo con el óvulo o en la zona de unión entre placenta y chalaza, mientras que en otras se extienden por toda la semilla.
Para favorecer la dispersión, algunas semillas poseen modificaciones de sus cubiertas tales como espinas, alas o paracaídas. Otro mecanismo de dispersión es cuando los animales las ingieren pero no las digieren, por ejemplo, cuando comen los frutos.
La semilla supuso un gran avance para el paso de un medio acuático a otro terrestre. Una de las grandes ventajas fue la capacidad para estar en estado de dormancia entre el momento de su liberación y el de la germinación. La dormancia es un estado de la semilla en el cual la tasa metabólica, o tasa de reacciones químicas dentro de la célula, disminuye, por lo que necesita muy poca energía, oxígeno o agua. Una combinación de factores externos como luz, agua, temperatura y sustancias químicas pueden terminar con este periodo. Esto permitió iniciar el crecimiento bajo condiciones favorables.
Este estado de quietud se consigue gracias a barreras biológicas creadas por la propia semilla, sus cubiertas, pero también a una detención de los procesos fisiológicos en el embrión y otros tejidos de la semilla. La dormancia, o más bien, la salida de la dormancia puede estar controlada por el embrión, por el endospermo, por la cubierta o por una combinación de estos. Al contrario que los animales, los embriones de las plantas pueden permanecer durmientes, permitiendo que una generación de semillas pueda producir plantas nuevas durante varios años. Se han encontrado semillas de más de 2000 años que han sido capaces de germinar y dar una planta adulta.
Las cubiertas de las semillas impiden que se dispersen los inhibidores del crecimiento, que en la mayoría de las plantas es principalmente el ácido abcísico, una hormona vegetal. Por otro lado, el ácido giberélico promueve la germinación. En Arabidopsis, la relación en la semilla de ácido abicísico/ácido giberélico es la que condiciona la salida o no de este periodo de dormancia.
Las semillas de una misma especie no abandonan la dormancia al mismo tiempo; incluso en condiciones favorables hay semillas que no iniciarán la germinación. En ocasiones, hay señales como el fuego, aparentemente no favorables, que facilitan la salida de la dormancia. Hay una estrategia de la mejor apuesta: algunas semillas saldrán de la dormancia bajo ciertas señales favorables, pero no lo harán todas. Las que no germinen tendrán que esperar a otra situación favorable. Otra estrategia es producir semillas morfológicamente diferentes o químicamente diferentes. Se ha demostrado que incluso semillas genómicamente idénticas tienen variabilidad en los tiempos de germinación, la cual puede ser generada por procesos estocásticos que generan un “ruido transcripcional”, es decir, la expresión un tanto aleatoria de genes.
Todas estas estrategias van encaminadas a repartir la germinación de una población de semillas en el tiempo y, así, como población, posibilitar la aparición de plantas en distintas condiciones y a distintos tiempos. Se ha propuesto que cada semilla tiene una relación distinta de ácido abcísico/ácido giberélico y que esto determina las condiciones de salida de la dormancia. El endospermo y el embrión de la semilla producen ácido abcísico, y el endospermo parece ser un elemento clave en el valor de esta relación entre hormonas. Sin embargo, la decisión de iniciar la germinación la toman células de la radícula del embrión.
La Germinación
La germinación es el proceso mediante el cual el embrión de la semilla se activa, se desarrolla y crece hasta la emisión de una radícula. La germinación se inicia normalmente con una entrada de agua a través del micropilo, residuo del tubo polínico producido durante la fecundación. Estas cubiertas son normalmente impermeables al agua por su cutícula con contenido en ceras.
Cuando se elimina la impermeabilidad por los elementos, hongos o patógenos, o por el paso por el tracto de los animales, el agua entra, así como el oxígeno, y los inhibidores son neutralizados. Se produce entonces un aumento del tamaño de las células del embrión, para lo que tienen que relajar sus paredes celulares. La división celular no es frecuente en este primer paso de la germinación. En Arabidopsis, la extensión celular se da sobre todo en la región del hipocótilo próxima a la radícula y en la zona de transición. Esta expansión lleva a la ruptura de la testa y es tanto longitudinal como radial, lo que crea presión en todas las direcciones. Le sigue una activación del metabolismo y una nueva fase de incorporación de agua acompañada de la emisión de la radícula, donde se considera que la germinación se ha completado.
Durante la germinación, el embrión emplea el material de reserva almacenado en el endospermo o en los cotiledones. La germinación ocurre en condiciones determinadas, las cuales dependen de la especie de planta. El endospermo se tiene que debilitar para permitir la germinación, normalmente por una señal del embrión y una producción de hormonas, sobre todo las giberelinas. En las plantas dicotiledóneas, es la radícula la primera estructura en crecer. Esto ocurre a la vez que el tejido provascular se transforma en xilema y floema funcionales. Posteriormente se desarrolla el epicótilo.
¿Qué Es La Germinación De Semillas? | Ciencia Para Niños | Vídeo educativo | Educativos Para Niños
tags: #embrion #definicion #plantas