La reproducción de los seres vivos y la transmisión de sus características hereditarias son procesos fundamentales que dependen íntimamente de los cromosomas y de la división celular. La forma en que las células se dividen y cómo los organismos se reproducen, ya sea de forma sexual o asexual, varía significativamente entre los distintos reinos de la vida, como las plantas, los animales y los hongos, cada uno con sus estrategias adaptativas.
División Celular: Mitosis y Cromosomas
Concepto General de la Fase M
La fase M es el periodo del ciclo celular donde ocurre la división de una célula madre en dos células hijas. Esta fase incluye una serie de procesos paralelos que buscan repartir de forma generalmente equitativa los componentes celulares, sintetizados en fases previas del ciclo. Estos componentes incluyen el ADN, duplicado en la fase S, y los elementos citoplasmáticos, producidos en las fases G1, S y G2. Durante la mitosis, se forman los cromosomas y se distribuyen entre las dos células hijas. Las etapas de la mitosis están ligadas a los eventos del ADN: su compactación, la formación y segregación de los cromosomas, y finalmente su descondensación en cromatina.
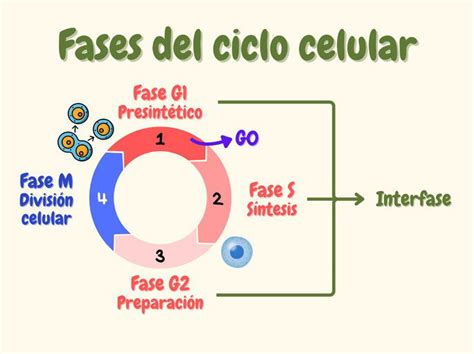
Fases de la Mitosis
La mitosis implica un cambio drástico en las células, incluyendo la formación del huso mitótico, una estructura de microtúbulos y cromosomas. En las células animales, los centrosomas se localizan en los polos de este huso, mientras que las plantas carecen de centrosomas pero también forman huso mitótico. Existen dos tipos de mitosis: abierta y cerrada. La mitosis abierta, común en animales y plantas, implica la desorganización de la envoltura nuclear para la formación del huso. Por el contrario, en la mitosis cerrada, el huso mitótico se forma dentro del núcleo, la envoltura nuclear no se rompe, pero se estrangula para dar origen a los dos núcleos hijos, evitando el contacto del citoplasma con los cromosomas. Algunas especies presentan formas intermedias donde la envoltura nuclear se conserva parcialmente o el huso se forma en el citoplasma sin ruptura de la envoltura.
Profase y Prometafase
La profase es la primera etapa de la mitosis. Comienza con la condensación del ADN, haciendo visibles las cromátidas, y la desaparición del nucléolo. Esta condensación es favorecida por la fosforilación de las histonas. En el citoplasma, se desorganizan parcialmente los filamentos del citoesqueleto y la célula pierde adhesividad, adquiriendo una forma redondeada, característica de las células en mitosis. Hacia el final de la fase S, las células animales duplican su centrosoma. Al inicio de la profase, los centrosomas se desplazan a polos opuestos del citoplasma, guiados por proteínas motoras y microtúbulos, donde nuclean y organizan el huso mitótico. Los orgánulos como el retículo endoplasmático y el aparato de Golgi se fragmentan, y el tráfico vesicular disminuye drásticamente. La envoltura nuclear permanece intacta. En el aparato de Golgi, una mayor fosforilación de GRASP y golginas durante la profase inhibe los mecanismos de anclaje de vesículas.
Algunos autores distinguen la prometafase, una fase al final de la profase, caracterizada por la fosforilación de las proteínas de la lámina nuclear, lo que desorganiza la envoltura nuclear en pequeñas vesículas, permitiendo el acceso de los microtúbulos a las cromátidas. Las cromátidas se compactan progresivamente en cromosomas típicos. Los microtúbulos establecen uniones con los cinetocoros, estructuras proteicas ensambladas en las regiones centroméricas de los cromosomas que median la interacción entre cromosomas y microtúbulos del huso. Cada cromosoma tiene dos cinetocoros orientados en direcciones opuestas, permitiendo que ambos polos del huso emitan microtúbulos que contactan el mismo cromosoma. Los microtúbulos que conectan con los cinetocoros se llaman cinetocóricos (20 a 40 en humanos, uno en levaduras). Otros microtúbulos, denominados polares o interpolares, parten de los polos del huso e interaccionan entre sí para estabilizarse. Finalmente, los microtúbulos astrales nuclean en los polos del huso hacia la membrana plasmática cercana.
Metafase: Regulación y Punto de Control
Al final de la profase (o prometafase), las cromátidas hermanas de cada cromosoma están unidas entre sí y a los microtúbulos cinetocóricos del huso. Los cromosomas son desplazados hacia el centro del huso, formando la placa ecuatorial, que define la metafase. Este movimiento es resultado del acortamiento y alargamiento de los microtúbulos y la acción de proteínas motoras. Durante este periodo, los cromosomas se posicionan en la placa ecuatorial, aunque a veces se desplazan temporalmente fuera de ella.
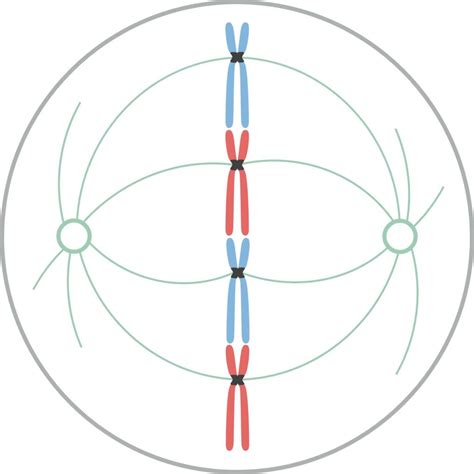
La correcta unión de los microtúbulos a los cinetocoros es crucial. Un reparto desigual de cromátidas podría conducir a aneuploidías, causando inviabilidad celular o patogenicidad. Para prevenirlo, la metafase cuenta con el punto de control del ensamblado del huso mitótico (SAC), que asegura que todos los cinetocoros estén conectados a un número apropiado de microtúbulos. Si un cinetocoro no está conectado, emite una señal que inhibe la segregación de cromátidas y la entrada en la anafase. El SAC está activo desde la profase hasta la metafase. Inhibe la actividad de la ubiquitina ligasa E3, conocida como APC/C, que normalmente se activa por cdc20 y marca las proteínas securina y ciclina B (inhibidoras de la anafase) para su degradación. Si hay cinetocoros desconectados, liberan una molécula que inhibe APC-cdc20, deteniendo el avance de la mitosis. La señal inhibitoria es proporcional al número de cinetocoros desunidos. Proteínas que componen este inhibidor se agregan al cinetocoro durante la profase y son eliminadas, por ejemplo, por dineínas, cuando el microtúbulo hace contacto.
La REGULACIÓN del Ciclo Celular
Errores en la mitosis causados por daños en el ADN (por ejemplo, drogas anticáncer) pueden activar el SAC de forma prolongada. Si una célula permanece mucho tiempo en prometafase, puede morir por apoptosis. Existe evidencia de que las células miden la duración de la mitosis, y si es demasiado larga, el ciclo celular se detendrá en G1 y podría llevar a apoptosis, lo que sugiere una "memoria" en G1 de los eventos de la fase M.
Anafase
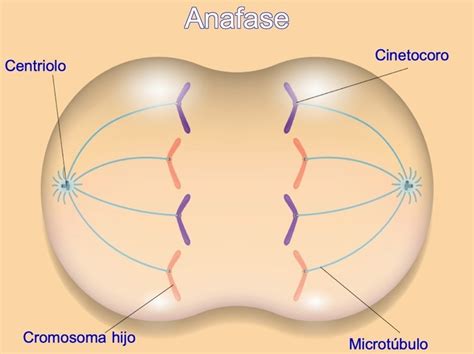
La anafase comienza con la ruptura de las conexiones entre cromátidas hermanas a nivel del centrómero, gracias a proteasas. Cada cromátida es arrastrada hacia uno de los centrosomas por los microtúbulos del huso a una velocidad de aproximadamente 1 µm por minuto. Se distinguen dos etapas:
- Anafase A: Los microtúbulos cinetocóricos se acortan por despolimerización en ambos extremos.
- Anafase B: Los centrosomas se separan entre sí, empujados por la elongación de los microtúbulos polares, con la fuerza aportada por proteínas motoras. Otras proteínas motoras asociadas a los microtúbulos del áster tiran de los centrosomas hacia la superficie celular, contribuyendo a la separación.
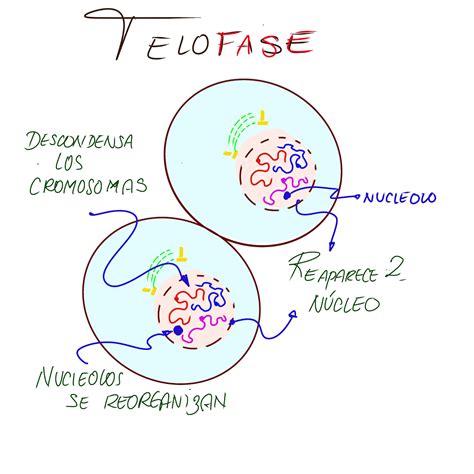
Telofase
Durante la telofase, la etapa final de la mitosis, la envoltura nuclear se reorganiza alrededor de cada conjunto de cromátidas que han migrado hacia los centrosomas, formando dos núcleos hijos. Este proceso implica la desfosforilación de las proteínas de la lámina nuclear. También se forman los poros nucleares y las cromátidas comienzan a descondensarse. Al finalizar la mitosis, una fuerte desfosforilación revierte la célula a su estado de G1.
Citocinesis: Separación del Citoplasma
La citocinesis es la etapa final del ciclo celular, donde el citoplasma de la célula madre se divide en dos partes, formando las células hijas. Esta separación ocurre después de la segregación completa de los cromosomas para evitar la formación de células con ploidías incorrectas. La citocinesis presenta diferencias notables entre animales, plantas y hongos.
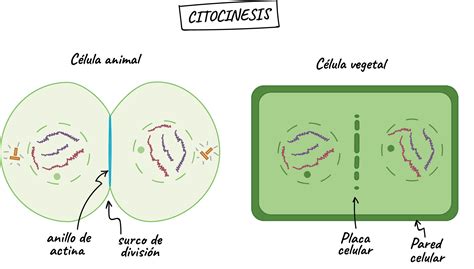
Citocinesis en Células Animales
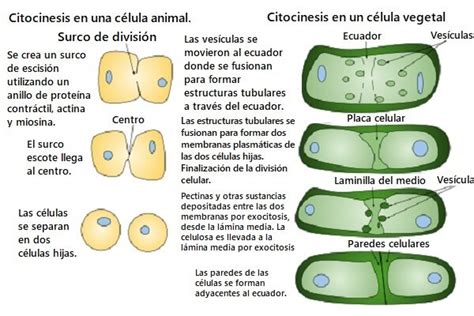
En células animales, el plano de división está determinado por la orientación del huso mitótico. El primer signo de citocinesis es la formación de un surco de escisión en la superficie celular, perpendicular al huso mitótico y situado en una posición ecuatorial. La posición del surco puede variar en divisiones asimétricas, resultando en células hijas de diferente tamaño. El surco de escisión es producido por la acción conjunta de filamentos de actina y la proteína motora miosina, que forman el anillo de escisión. Este anillo se ensambla al final de la anafase y su contracción, similar a la muscular, provoca un estrangulamiento. Es una estructura transitoria que desaparece tras completar la citocinesis, que también requiere la eliminación de los restos del huso mitótico y la ruptura y sellado de las membranas plasmáticas.
El tráfico vesicular también juega un papel crucial en la finalización de la citocinesis en células animales, al igual que en vegetales. Se requiere la adición de membrana a la membrana plasmática antes, durante y después de la formación del anillo de escisión para crear las nuevas células hijas, manteniendo un balance entre endocitosis y exocitosis. Las proteínas en torno al anillo de constricción capturan vesículas del Golgi y de los endosomas. En las células animales, al cerrarse el anillo, queda atrapada una estructura electrondensa llamada cuerpo medio, compuesta por microtúbulos y actina. El paso final es la fusión de las membranas del surco, con la participación del complejo ESCRT-III. Además, la remodelación de la matriz extracelular es necesaria en células animales.
Citocinesis en Células Vegetales y Hongos
En células vegetales y hongos, la citocinesis difiere debido a la presencia de la pared celular. En las plantas, la separación de las células hijas no se produce por un anillo contráctil, sino por la formación de una nueva pared celular dentro de la célula madre, que eventualmente las separará. La formación de esta nueva pared está mediada por el fragmoplasto, compuesto inicialmente por microtúbulos polares del huso mitótico y vesículas del aparato de Golgi. Estas vesículas son transportadas a la zona media, se fusionan para formar membrana, y su contenido constituye la lámina media de la futura pared celular. La nueva pared crece de manera centrífuga.
En los hongos, la citocinesis también es particular. En animales y plantas, la cariogamia (fusión de núcleos) suele seguir rápidamente a la plasmogamia (fusión celular). Sin embargo, en la mayoría de los hongos, los núcleos de distinto origen pueden coexistir en el citoplasma sin fusionarse, dando lugar a un micelio con dos tipos de núcleos (dicariótico) o un número indeterminado (heterocariosis). Por ejemplo, en un basidiomiceto típico, la cariogamia ocurre solo en los basidios, siendo el resto del micelio dicariótico. Esto permite que criaturas haploides como la mayoría de los hongos superiores, tengan varios alelos del mismo gen, complicando el estudio de su genética.
Reproducción y Variabilidad Genética
Reproducción Asexual vs. Sexual
Aunque no es indispensable para todos, la capacidad de generar descendencia a través del sexo es muy común en eucariontes: protistas, algas, plantas, animales y hongos. Es probable que el ancestro común de todos ellos ya lo hiciera. La reproducción sexual, a diferencia de la asexual, genera mayor variación genética. En ella, la meiosis y la recombinación mezclan aleatoriamente fragmentos de los genomas parentales, produciendo un genoma único en cada descendiente. En contraste, en la reproducción asexual, los nuevos individuos son casi copias idénticas del organismo de origen, con ligeras diferencias por mutaciones azarosas durante la duplicación del material genético en la mitosis.
Reproducción Sexual en Animales
Los seres humanos y muchos otros animales somos diploides, lo que significa que poseemos dos juegos de cromosomas en nuestras células. Cuando se forman las células sexuales (gametos), estas son haploides, portando un solo juego de cromosomas. La reproducción sexual inicia con la unión de dos gametos haploides (plasmogamia), seguida de la fusión de sus núcleos (cariogamia) para formar una célula diploide, el zigoto, que se desarrollará hasta convertirse en un adulto. En los seres humanos y muchos otros animales, el sexo está determinado por los genes de los cromosomas sexuales X e Y, que codifican la información para el desarrollo de órganos sexuales y diferencias morfológicas.
Particularidades de la Reproducción en Hongos
Los hongos han desarrollado la capacidad de aprovechar ambas modalidades reproductivas. La reproducción asexual es ventajosa en ambientes estables con alimento abundante, permitiendo la proliferación rápida de individuos idénticos. Un ejemplo es el hongo fitopatógeno Puccinia graminis, que produce millones de esporas asexuales en un campo de trigo. Sin embargo, en ambientes inhóspitos, la reproducción sexual puede ser más ventajosa, produciendo esporas de resistencia con individuos recombinantes.
La REGULACIÓN del Ciclo Celular
Haploidía y Ciclos de Vida
Los núcleos de plantas y animales son generalmente diploides, mientras que en los hongos son casi siempre haploides, es decir, tienen una sola copia de cada cromosoma. Esto implica ciclos de vida muy diferentes. Mientras que en animales, el ciclo comienza con la unión de gametos haploides para formar un cigoto diploide, en la mayoría de los hongos, el ciclo inicia con la producción de esporas haploides y recombinantes que germinan para formar un nuevo talo. Algunos hongos permanecen haploides casi toda su vida; otros, como las setas, fusionan sus hifas al germinar las esporas y forman un micelio dicariótico con dos núcleos haploides separados en cada célula, lo que les permite tener dos copias de cada cromosoma, aunque en núcleos distintos.
Determinación del Sexo: Loci y Múltiples Sexos
A diferencia de animales, en los hongos no hay cromosomas sexuales y la mayoría no presenta diferencias morfológicas entre lo que se reconocería como femenino y masculino. No obstante, la mayoría posee dos tipos de compatibilidad, es decir, dos sexos. En estos sistemas, el sexo está determinado por un locus (región específica del genoma) que puede contener uno o más genes distintos y complementarios en cada sexo. Las funciones de este locus incluyen la producción de feromonas para atraer parejas potenciales, su recepción, la meiosis y la formación de estructuras reproductivas.
En algunas especies de Agaricomycetes, el sexo está determinado por dos regiones (loci) distantes en el genoma, lo que permite la existencia de miles de sexos. Las hifas de estos hongos se atraen por feromonas codificadas en el primer locus (B); para la atracción, las feromonas deben ser diferentes. Una vez fusionadas las hifas, sus células se mezclan. Para que esta fusión sea exitosa, deben formar proteínas compatibles codificadas en el segundo locus (A), que también presenta muchos alelos. Para la reproducción, los individuos deben tener alelos diferentes en ambos loci. Dada la cantidad de alelos, especies como Schizophyllum commune pueden tener más de 20 mil combinaciones o sexos.
Estrategias Reproductivas Inusuales en Hongos
Algunos hongos presentan sistemas reproductivos muy inusuales. Por ejemplo, la levadura de cerveza (Saccharomyces cerevisiae) es un organismo unicelular que puede reproducirse sexual o asexualmente y cambia entre sus dos sexos posibles después de cada mitosis mediante edición del ADN. Esto asegura una población equilibrada de ambos sexos para la reproducción sexual. A pesar de que los apareamientos entre levaduras "hermanas" son frecuentes, la reproducción sexual sigue siendo ventajosa porque durante la meiosis ocurre un proceso de "limpieza" y reparación del ADN.
Las funciones del sexo van más allá de generar variación. El hongo fimícola Podospora anserina utiliza los ciclos sexuales para eliminar genes duplicados sin función aparente (genes egoístas) que pueden multiplicarse y causar mutaciones indeseables en el genoma.
Heterocariosis y Ciclo Parasexual
Algunos hongos han perdido la capacidad de reproducción sexual, pero han desarrollado sistemas para mantener o aumentar la variación genética. Muchos mohos, como Penicillium o Aspergillus, pueden formar hifas con núcleos haploides de dos o más individuos, fenómeno llamado heterocariosis. Esto ocurre como un efecto secundario de su sistema de autorreconocimiento, análogo al sistema inmunitario. Hifas que se reconocen como propias (con alelos idénticos en ciertos loci) se fusionan, y los genes de ambos núcleos se complementan, lo que es beneficioso en ambientes donde no sobrevivirían individualmente. La heterocariosis puede dar lugar a un ciclo parasexual, generando variación equivalente a la recombinación. Ocasionalmente, los dos núcleos diferentes se fusionan, formando núcleos diploides que, tras varias mitosis inestables, producen núcleos haploides con combinaciones aleatorias de los cromosomas originales. Estos núcleos recombinantes pueden dispersarse en esporas asexuales, colonizando nuevos ambientes.
Adaptación y Consecuencias para los Humanos
La Versatilidad Sexual de los Hongos y el Tamaño Poblacional
La alta capacidad de adaptación de los hongos se debe, en parte, a su versatilidad sexual y al enorme tamaño de muchas de sus poblaciones. La reproducción asexual genera millones de individuos casi idénticos con algunas mutaciones nuevas y diferentes, que ocurren de forma aleatoria e independiente en cada mitosis. La mayoría de las mutaciones no afectan la eficacia, muchas son perjudiciales y muy pocas son ventajosas. Sin embargo, en poblaciones más grandes, las posibilidades de tener mutaciones ventajosas en menos tiempo aumentan.
Aplicaciones Biotecnológicas y Resistencia a Antifúngicos
Los seres humanos han aprovechado este principio en la biotecnología. Por ejemplo, para la producción industrial de penicilina a partir de Penicillium chrysogenum, se usaron técnicas que aumentaron la tasa de mutación en miles de individuos experimentales, creando variantes que incrementaron la producción 15 mil veces. Sin embargo, esta misma capacidad adaptativa puede ser perjudicial. El uso indiscriminado de antifúngicos (azoles) en la agricultura intensiva está provocando resistencia, lo que representa un riesgo para la seguridad alimentaria. Además, muchos hongos del suelo pueden causar infecciones oportunistas en humanos, que solo pueden tratarse con azoles.
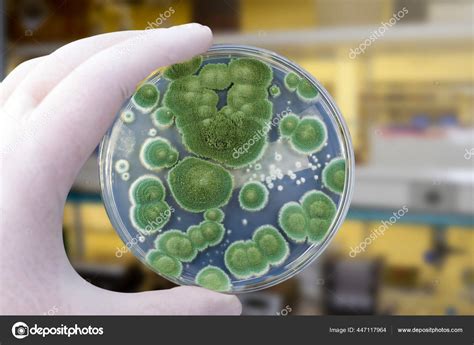
La variación genética de algunos hongos ha sido moldeada por los humanos incluso antes de comprender la herencia. La producción de cerveza industrial depende de linajes de levadura domesticados en Europa desde el siglo XVII, seleccionándose características como la gran capacidad para usar maltosa, la reducida producción de metabolitos de mal sabor, la pérdida de genes indispensables para la vida libre, y una casi nula reproducción sexual.
Importancia del Estudio de la Genética Fúngica
Conocer los sistemas genéticos de los hongos es vital para crear estrategias que mitiguen sus efectos negativos y, al mismo tiempo, aprovecharlos al máximo. Estudiarlos desde esta perspectiva nos proporciona herramientas para comprender mejor la naturaleza misma del sexo, su evolución y los alcances y limitaciones de la variación. Aunque la variabilidad genética es crucial, existen otras variaciones igualmente relevantes en la evolución, como las que ocurren durante el desarrollo individual hacia la vida adulta y las que implican la expresión de diferentes morfologías ante cambios ambientales sin alteraciones genéticas.
tags: #cromosomas #reproduccion #en #plantas #animales #y