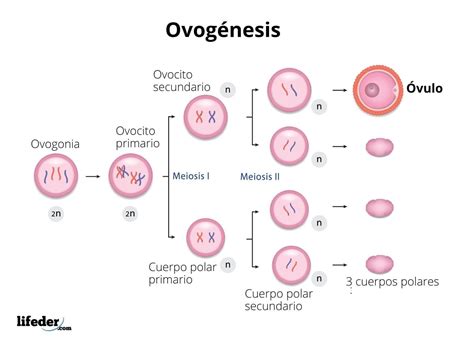
La ovogénesis se define como la formación de los gametos femeninos. Este es un proceso cíclico que sucede en el ovario, durante el cual las células germinales primordiales se transforman en ovocitos. La ovogénesis es un proceso complejo que involucra varias fases de desarrollo y arresto meiótico, esenciales para la correcta maduración del ovocito y su potencial para la fecundación.
Introducción a la Ovogénesis y el Bloqueo Meiótico
Los gonocitos, o células germinativas primordiales, migran a los ovarios del embrión. En esta fase, las ovogonias pueden seguir dos caminos: la mayoría sigue dividiéndose por mitosis, mientras que otras aumentan su tamaño al acumular mayor masa citoplasmática en su interior. En el quinto mes del desarrollo embrionario, se alcanza el número máximo de folículos primordiales, aproximadamente 7 millones.
Estos folículos primarios contienen en su interior a los ovocitos que bloquean su división meiótica en la fase de diploteno de la profase I. Este bloqueo se mantiene debido a la acción del inhibidor de maduración ovocitaria (OMI), secretado por las células foliculares que rodean al ovocito. Los ovocitos permanecen en este arresto meiótico durante un periodo prolongado, a menudo por décadas en la mujer.
El Ovocito de Xenopus laevis como Modelo de Estudio
El ovocito de Xenopus laevis (X. laevis) es un sistema de expresión heteróloga óptimo para el análisis de las vías de señalización celular. Su éxito como modelo se debe, en parte, a que sus ovocitos son las células individuales más grandes del reino animal, facilitando su manipulación y el microinyección de DNA o mRNA. Los ovocitos de X. laevis son de un mantenimiento relativamente simple y su manipulación es fácil y asequible, no requiriendo del sacrificio de las ranas para su obtención.
En X. laevis, los ovocitos permanecen en un arresto meiótico muy prolongado en profase I durante su desarrollo, similar al de los mamíferos, lo que los hace un modelo excelente para estudiar los mecanismos de mantenimiento y reanudación de la meiosis.
Mantenimiento del Bloqueo Meiótico en Profase I
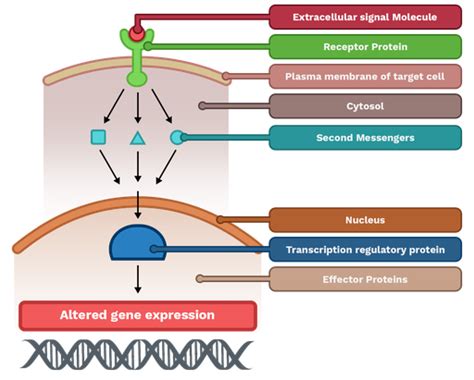
El principal regulador del arresto meiótico del ovocito es el adenosín monofosfato cíclico (AMPc). Niveles elevados de AMPc son cruciales para mantener el ovocito en profase I. La vía de señalización clave para mantener este arresto implica la activación de las proteínas Gαs, que, a su vez, activan a la adenilato ciclasa (AC). La AC cataliza la producción de AMPc, lo que lleva a la activación de la cinasa dependiente de AMPc (PKA), la cual fosforila proteínas clave que mantienen el ovocito en el estado de arresto meiótico en profase I.
Reanudación de la Meiosis I: Desactivación del Bloqueo
Una vez llegada la pubertad, en la mujer comienzan a producirse ciclos de ovulación. En cada uno de estos ciclos, varios ovocitos primarios reactivan su meiosis I, proceso esencial para la maduración final del ovocito. Esta reanudación es activada por hormonas, principalmente esteroides como la progesterona en el caso de Xenopus, que provocan el rompimiento de la vesícula germinal (GVBD).
Mecanismo de Acción de la Progesterona
La progesterona actúa mediante un receptor membranal acoplado a proteínas Gi. Este modelo propone que la progesterona funciona como un agonista que activa a una proteína Gi, que inhibe la AC por medio de sus subunidades Gαi. La inhibición de la AC reduce los niveles intracelulares de AMPc. Esta reducción de AMPc lleva a la inactivación de la PKA, lo que permite la activación del Factor Promotor de la Maduración (MPF).
Meiosis Paso a Paso
El Factor Promotor de la Maduración (MPF)
El MPF es un complejo proteico que controla la transición de la fase G2 a la meiosis I (profase I). Fue inicialmente denominado "maturation promoting factor" por Masui y Market en 1971. Está compuesto por una cinasa dependiente de ciclina (Cdc2) y una ciclina (ciclina B). En los ovocitos en arresto, el MPF se encuentra en un estado inactivo (pre-MPF) debido a la fosforilación de sus residuos de tirosina (Y15) y treonina (T14) en la cinasa Cdc2.
La progesterona estimula la desfosforilación de la Y15 y T14, lo que activa el MPF y promueve la maduración del ovocito. Existen dos vías principales por las cuales el pre-MPF se activa:
- La proteína CDC25 desfosforila directamente la Cdc2 del MPF, activándolo.
- Un mRNA materno almacenado codifica la proteína Mos, que activa la vía MEK-MAPK-90Rsk. Esta vía, a su vez, activa indirectamente al MPF. La progesterona en los ovocitos de X. laevis también aumenta la expresión de la proteína Mos.
La activación del MPF lleva a una serie de eventos cruciales para la reanudación de la meiosis I, incluyendo la fosforilación de proteínas nucleares, el rompimiento de la envoltura nuclear (GVBD), la condensación de los cromosomas y la formación del huso meiótico. Tras esta primera división, el ovocito primario forma dos células hijas diploides de tamaño diferente: una célula de mayor tamaño que es el ovocito secundario y una de menor tamaño, el primer corpúsculo polar, que queda retenida en el espacio perivitelino.
El Segundo Bloqueo Meiótico: Metafase II
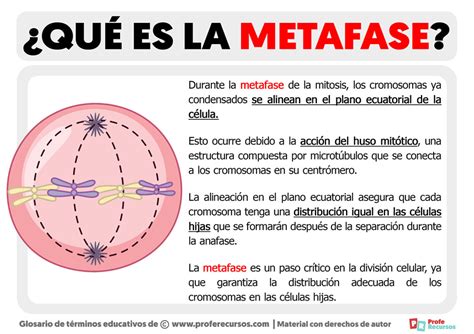
La ovulación se produce con el ovocito en metafase II, lo que representa un segundo punto de arresto meiótico. Este arresto es mantenido por las acciones combinadas de las proteínas c-mos y la cinasa 2 dependiente de ciclina (Cdc2). Este complejo, conocido como el Factor Citostático (CSF), bloquea el ciclo celular en metafase hasta que se produce la fecundación.
Reanudación de la Meiosis II: Tras la Fecundación
El ovocito secundario solo acaba de madurar si es fecundado. El ciclo celular que se detiene en metafase II se reanuda en el momento de la fecundación. Este proceso es desencadenado por el incremento de iones de calcio libre en el citoplasma después de la fecundación. El calcio activa la proteína calmodulina, que a su vez activa a la calpaína II, una proteasa dependiente de calcio. La calpaína II degrada la proteína c-mos, lo que desactiva el CSF. La degradación de c-mos promueve la división celular, y así, se completa la segunda división meiótica, liberando el segundo corpúsculo polar y dando lugar al óvulo maduro, listo para fusionarse con el espermatozoide.
Canales Iónicos: El Canal GIRK5 en la Maduración Ovocitaria
Los canales iónicos de potasio son proteínas de membrana que permiten el flujo de iones potasio a través de la membrana celular a favor de su gradiente electroquímico, modulando el potencial de membrana y la excitabilidad celular.
Canales GIRK (Kir3)
Los canales de potasio del tipo rectificador entrante (Kir), específicamente la subfamilia Kir 3 (GIRK, del inglés "G protein-coupled inwardly rectifying potassium channel"), son importantes en la neurotransmisión inhibitoria y en la regulación de la actividad celular. Se caracterizan por ser activados por subunidades Gβγ de proteínas G heterotriméricas.
GIRK5 en Ovocitos de Xenopus laevis
El canal de potasio GIRK5 es un canal endógeno localizado en los ovocitos de X. laevis. Forma homomultímeros funcionales que son activados por una poza endógena de subunidades Gβγ. Los canales GIRK5 se activan por ligandos de receptores metabotrópicos acoplados a proteínas Gi/G0. Estas proteínas Gi heterotriméricas constan de tres subunidades: alfa (αi), beta (β) y gamma (γ).
La expresión funcional del canal GIRK5 está regulada por la fosforilación de un residuo de tirosina 16 (Y16) en el amino terminal. Esta regulación es mediada por cinasas de tirosinas (PTKs) endógenas. La mutación de la tirosina 16 a alanina (GIRK5-Y16A) permite que el canal se exprese en forma funcional en la membrana plasmática de los ovocitos de X. laevis. Esto sugiere que la fosforilación de la tirosina 16 es importante para la regulación de la expresión funcional del canal GIRK5 y su participación en el proceso de maduración del ovocito.