La formación del gameto femenino en la mayoría de las hembras de mamíferos, incluida la especie humana, comienza en la etapa embrio-fetal. En esta fase temprana, se determina el número definitivo de células germinales del individuo. Durante la etapa fértil, el ovario responde a la secreción cíclica de gonadotropinas (LH y FSH), generando y liberando periódicamente ovocitos aptos para ser fecundados, lo que constituye su función citogénica. Paralelamente, el ovario secreta hormonas esteroidales, peptídicas y factores de crecimiento, cumpliendo una función endocrina. La obtención normal del gameto femenino depende intrínsecamente del desarrollo adecuado del folículo al que pertenece.
Los cambios fisiológicos ováricos con la edad, como la disminución progresiva de la fertilidad en mujeres mayores de 35 años, se deben esencialmente a una depleción de la población folicular. Cuando esta población desciende por debajo de 1000 folículos, el ovario pierde su capacidad de ciclar normalmente, lo que conlleva una disminución en los niveles de esteroides circulantes y sus efectos secundarios. Asociado a esta reducción folicular, se producen cambios en la calidad ovocitaria, manifestándose en un aumento de aneuploidías, disfunciones mitocondriales, alteraciones de la microcirculación y una disminución de la capacidad defensiva contra las especies reactivas del oxígeno (ROS).
Desarrollo Ovárico Embrionario y el Ovocito Primario (Ovocito I)
Durante la etapa embrionaria, las células germinales primordiales (CGP) inician su desarrollo entre la 5.ª y 6.ª semana de gestación. Cien a doscientas CGP colonizan la gónada embrionaria y, a través de mitosis, su número aumenta y se diferencian en ovogonias. Este proceso ocurre gradualmente en el ovario fetal. Las ovogonias, células diploides (2n, 2c), se multiplican por mitosis, alcanzando alrededor de 7 millones en el ovario fetal a las 20 semanas de gestación. Este proceso de división mitótica se considera la fase proliferativa, cuyo propósito es crear un reservorio celular durante la vida fetal. De esta división se obtienen dos células hijas iguales a la madre por cada ovogonia.
🌐 División celular: MITOSIS 🔬 | FASES | [Fácil y Rápido] | BIOLOGÍA |
Como células germinales, las ovogonias tienen la capacidad de experimentar meiosis. Así, entre la octava y la decimotercera semana de gestación, las ovogonias ingresan paulatinamente a la primera profase meiótica, transformándose en ovocitos I. La primera meiosis se detiene en el estado de dictioteno, hacia finales de la profase I, sin progresar a la metafase I. En este estado, el núcleo del ovocito se denomina vesícula germinativa debido a sus características morfológicas.
En la especie humana, este primer reposo meiótico puede durar más de 45 años. Aunque la actividad meiótica del ovocito I está detenida durante este periodo, la actividad transcripcional y, en menor grado, la traduccional, permanecen activas, lo que permite la supervivencia de la célula germinal. De manera coincidente con la generación de ovocitos I en profase I, células somáticas del estroma ovárico rodean a cada ovocito I, iniciando la formación de los folículos primordiales.
Estas células somáticas corresponden a las células pre-granulosas, que secretan una matriz extracelular formando la lámina basal que rodea al folículo primordial. La interacción entre las células somáticas y el ovocito es posible gracias a que las células de pre-granulosa expresan el Stem Cell Factor (SCF), un ligando para una proteína receptora específica en la membrana plasmática del ovocito. Si esta interacción molecular específica no ocurre, no se forman folículos primordiales y el ovocito muere por apoptosis.
Hacia finales de la gestación, todos los ovocitos I presentes en el ovario fetal están detenidos en profase I de la meiosis, formando parte del folículo primordial. Al nacer, la población de folículos primordiales en ambos ovarios es de alrededor de 2 millones, constituyendo el potencial reproductivo de la mujer. A diferencia de anfibios y aves, en mamíferos no se produce vitelo en el ovocito durante la ovogénesis.
Foliculogénesis: Del Folículo Primordial al de Graaf
La foliculogénesis es el proceso de crecimiento que experimenta un folículo desde el momento en que deja la población de reserva (folículos primordiales) hasta su ovulación o atresia. En un ovario postpuberal, los folículos pueden encontrarse en distintos estados de desarrollo: primordiales, primarios, secundarios, terciarios (antrales) y ovulatorios (de Graaf) o atrésicos.
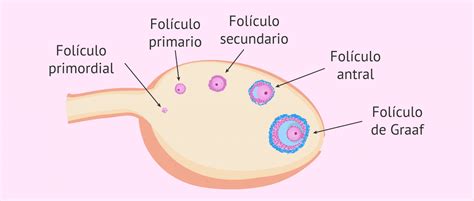
El crecimiento y desarrollo de los folículos desde el estado primario hasta el maduro implica la proliferación de las células de la granulosa (CG) y el crecimiento del ovocito I, lo que resulta en un aumento del diámetro folicular. El ovocito crece fundamentalmente hasta la etapa de folículo primario tardío o secundario inicial, aumentando su diámetro de 30μm a 120μm. Durante este periodo, se forma la zona pelúcida, un material glicoproteico de origen ovocitario que rodea completamente al ovocito. Asimismo, el número de mitocondrias, ribosomas y otros orgánulos del ovocito aumenta considerablemente, contribuyendo al tamaño de la célula germinal.
En esta etapa, las células de la granulosa inician la expresión de receptores para FSH. Al final del estado secundario, ocurre una migración de células mesenquimáticas del estroma ovárico, dando origen a las células de la teca interna (TI) y externa (TE), lo que marca el inicio de la formación del folículo terciario. Simultáneamente, el folículo terciario comienza a formar el antro, una cavidad llena de fluido folicular que se acumula entre las CG. Este fluido está compuesto por productos de secreción de las CG y transudado de los capilares sanguíneos que irrigan las células tecales.
También en esta etapa, las células tecales, las CG entre sí, y las CG de la corona (las más cercanas al ovocito) forman uniones comunicantes o gap, lo que implica que están acopladas eléctrica y metabólicamente. Esto permite una sincronización funcional en respuesta a la estimulación por diversos factores, como los factores de crecimiento transformante beta (TGF-β). Se ha demostrado que la conexina 43 es esencial para la formación de estas uniones GAP entre el ovocito y las células del cúmulus.
Los folículos terciarios adquieren una gran capacidad para sintetizar andrógenos a nivel de las células de la teca interna, bajo el estímulo de la LH. Esto es posible porque en esta etapa de la foliculogénesis se expresan receptores para LH en dichas células. Por otro lado, el folículo se hace sensible a la acción de la FSH, cuyos receptores se expresan en las CG. La FSH activa la proteína quinasa A (PKA), que a su vez fosforila diversos sustratos, como la enzima citocromo P450 aromatasa, encargada de transformar andrógenos en estrógenos. La actividad y expresión de la aromatasa aumentan, incrementando gradualmente la producción de estrógenos.
La foliculogénesis se divide en una etapa basal (independiente de gonadotropinas) y una etapa tónica (dependiente de gonadotropinas como FSH y LH). Factores como el GDF9, a través de SMAD 2 y 3, son responsables de la transición de folículo primordial a primario, mientras que la AMH producida por folículos terciarios puede inhibir esta transición en folículos primordiales. El nivel de vascularización está directamente relacionado con el folículo dominante; un folículo con mayor flujo sanguíneo es el que será ovulado.
El Ovocito Secundario (Ovocito II): Reinicio Meiótico y Ovulación
Los ovocitos, tras su formación como ovocitos primarios, detienen el proceso de meiosis en dos ocasiones principales. La primera pausa ocurre durante la profase I de la primera división meiótica, como se mencionó, y no termina hasta el inicio de la pubertad y la estimulación hormonal. Un marcador clave del reinicio de la división meiótica es la ruptura de la vesícula germinal (GVBD), que libera el material genético para su posterior anclaje, alineación y distribución.
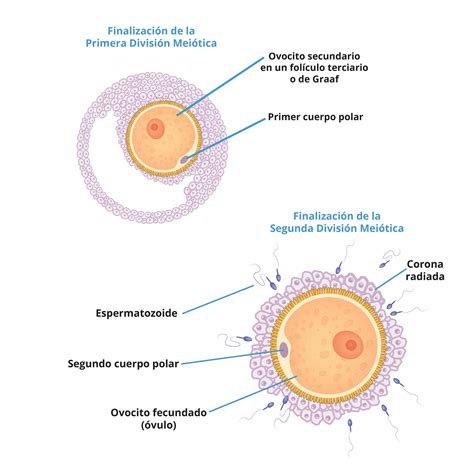
Estimulado por el pico de la hormona luteinizante (LH) liberada por la hipófisis, el ovocito reinicia la meiosis y completa su primera división meiótica. En esta primera división, un ovocito I diploide (2n, 4c, tras replicación de ADN) se divide en dos células haploides (1n, 2c): un ovocito de segundo orden (Ovocito II) y un primer corpúsculo polar. Este corpúsculo polar, que se atrofia, contiene la mitad del material genético pero casi nada de citoplasma, asegurando que el ovocito II retenga la mayor parte de los nutrientes y organelos.
🌐 División celular: MITOSIS 🔬 | FASES | [Fácil y Rápido] | BIOLOGÍA |
El ovocito secundario, ahora haploide, entra en la segunda división meiótica pero se detiene en metafase II. Esta segunda pausa meiótica se mantiene hasta que ocurre la fecundación. Al alcanzar el estado preovulatorio, las células de la granulosa adquieren receptores de LH, y el pico de LH induce la ovulación. En este momento, los vasos sanguíneos de la teca sufren vasoconstricción, generando un estigma por el cual se libera el ovocito. Cuando el folículo se rompe y el ovocito es expulsado del ovario, ya ha terminado la meiosis I y se expulsa un Ovocito II, rodeado de la zona pelúcida y las células foliculares más cercanas, conocidas como la corona radiata.
🌐 División celular: MITOSIS 🔬 | FASES | [Fácil y Rápido] | BIOLOGÍA |
A diferencia de la espermatogénesis, donde se obtienen cuatro espermatozoides haploides funcionales de una célula germinal, en la ovogénesis el ovocito resulta en una sola célula funcional con la mitad del material genético, debido a la expulsión de material genético en los corpúsculos polares. Este proceso de reducción y expulsión se conoce como maduración nuclear.
Morfología y Función de la Zona Pelúcida (ZP)
La Zona Pelúcida (ZP) es una matriz glicoproteica translúcida que rodea al ovocito de los mamíferos y se mantiene en el embrión preimplantacional hasta el estadio de blastocisto, aunque su estructura y funcionalidad varían. En humanos, está compuesta por cuatro glicoproteínas: ZP1, ZP2, ZP3 y ZP4. Estas presentan un patrón de glicosilación especie-específico, fundamental para el reconocimiento gamético, la inducción de la reacción acrosómica del espermatozoide y el bloqueo de la polispermia.
Además, durante el desarrollo embrionario preimplantacional, la ZP es esencial para la protección del embrión, la comunicación materno-cigótica, la prevención de la gemelaridad monocigótica (mediante la unión de los blastómeros) y la diferenciación de los blastómeros a trofoblasto (hipótesis de polarización). También juega un papel importante en la eclosión o hatching embrionario, un proceso que depende tanto de la acción mecánica y enzimática del blastocisto como de la actividad lítica uterina.
La disposición de los filamentos de la ZP le confiere una propiedad óptica denominada birrefringencia, lo que permite su análisis por microscopía de polarización (PolScope) y la definición de su estructura trilaminar. En el ovocito, los filamentos de la capa interna (capa 1) están orientados radialmente, mientras que los de la capa externa (capa 3) están orientados tangencialmente, lo que las hace muy birrefringentes. La capa intermedia (capa 2) presenta una birrefringencia mínima, sugiriendo una orientación aleatoria de los filamentos. Tanto en el ovocito como en el embrión temprano, la capa más interna es la más gruesa y la que mayor retardancia exhibe. La disminución del grosor general de la ZP en los embriones se debe, sobre todo, a la disminución de la capa más externa.

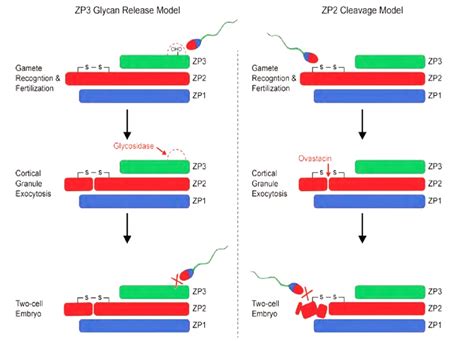
Las glicoproteínas de la ZP parecen ser secretadas coordinadamente por el ovocito durante la foliculogénesis, aunque en algunas especies (incluida la humana) las células de la granulosa también contribuyen a su producción. En el modelo murino, ZP2 y ZP3 interactúan formando unidades heterodiméricas que se disponen periódicamente en filamentos largos, interconectados por ZP1. Esta capa separa el oolema de la corona radiata.
En cuanto al reconocimiento espermatozoide-ovocito, el modelo clásico de liberación de glicanos propone que los O-glicanos de ZP3 actúan como ligandos para un receptor de superficie espermático, induciendo la reacción acrosómica. Tras la fecundación, los glicanos de la ZP3 serían eliminados, evitando la polispermia. Sin embargo, investigaciones recientes sugieren un modelo de escisión de ZP2, donde el ligando principal para los espermatozoides humanos es un dominio N-terminal de ZP2. Es posible que ambos modelos sean complementarios, con una unión inicial a ZP3 que induce la reacción acrosómica, seguida de la unión de espermatozoides ya reaccionados a ZP2.
| Grosor (µm) | Retardancia (nm) | |
|---|---|---|
| Ovocitos Inmaduros | 16.8 ± 1.2 | 25.3 ± 2.1 |
| Ovocitos Madurosa | 17.1 ± 1.1 | 26.5 ± 1.8 |
| Embriones Día 3b | 15.4 ± 1.0 | 22.9 ± 1.5 |
| Nota: Los valores son la media ± DE (one-tailed t-test; P < 0.05). a Diferencia estadísticamente significativa entre ovocitos inmaduros y maduros. b Disminución estadísticamente significativa entre ovocitos maduros y embriones en día 3. (Adaptada de Pelletier et al, 2004) | ||
Fecundación, Meiosis II e Implantación
La ovogénesis se completa cuando ocurre la fecundación. En ese momento, el ovocito II reanuda y finaliza la segunda división meiótica, dando lugar a un óvulo y un segundo corpúsculo polar. El óvulo no necesita diferenciación adicional y está listo para la fusión con el espermatozoide. La fecundación en humanos ocurre generalmente en la ampolla de las trompas de Falopio.
Tras la ovulación, los restos del folículo en el ovario se transforman en el cuerpo lúteo (cuerpo amarillo). Las células foliculares que lo forman secretan progesterona y estrógenos. Si no hay fecundación, el cuerpo lúteo degenera y desaparece, dando lugar al cuerpo blanco (corpus albicans), y el útero se descama, produciéndose la menstruación. Sin embargo, si ha habido fecundación, el cuerpo lúteo crece en tamaño y se mantiene, produciendo progestágenos y estrógenos, principalmente durante las primeras semanas del embarazo. El embrión, una vez implantado en el útero, comienza a secretar gonadotropina coriónica humana (hCG) para mantener el cuerpo lúteo activo durante la gestación.
Los óvulos liberados durante la ovulación son recogidos por las trompas de Falopio y conducidos hasta la cavidad del útero. El óvulo no puede moverse por sí mismo; su desplazamiento es provocado por los movimientos peristálticos de las trompas y la actividad de las células ciliadas del epitelio de la mucosa. El útero es el lugar donde ocurre la mayor parte del desarrollo embrionario. El embrión se implanta en el endometrio, formando la placenta con componentes de origen embrionario (corion) y materno (decidua).
El embrión está revestido por la ZP hasta poco antes de la implantación. Por lo tanto, todas las señales materno-embrionarias deben atravesarla. Las proteínas secretadas por el oviducto y el útero, así como las embrionarias, se incorporan a la ZP, cambiando sus propiedades morfológicas y bioquímicas. La eclosión de los blastocistos tardíos se debe a su acción mecánica (contracciones y reexpansiones) y a la acción de proteinasas como la tripsina (Strypsin) y las catepsinas. Además de la fuerza mecánica y las enzimas embrionarias, existe una contribución uterina al escape de la ZP mediante la producción de lisinas uterinas que ayudan a la disolución de la matriz glicoproteica, especialmente en el proceso in vivo.