El concepto de músculo estriado comprende dos tipos de tejido: el músculo esquelético y el músculo cardíaco. El músculo esquelético es el tejido del que están hechos los músculos unidos a los huesos, lo que nos permite el movimiento. Sus características fisiológicas principales son la excitabilidad y la contractilidad. Microscópicamente, tanto la musculatura esquelética como la cardíaca presentan una apariencia estriada debido a sus miofibrillas densamente empaquetadas, de ahí su denominación como tejido muscular estriado.
Para describir las estructuras celulares relacionadas con la musculatura esquelética se usan términos especiales que a menudo empiezan con los prefijos "mio-" o "sarco-". Por ejemplo, el citoplasma de una célula muscular es denominado sarcoplasma, la membrana plasmática se denomina sarcolema y el retículo endoplasmático se denomina retículo sarcoplasmático.
Orígenes Embrionarios Específicos del Músculo Estriado
Los distintos orígenes embriológicos del tejido muscular estriado lo hacen un tejido interesante. La musculatura de la cara, por ejemplo, proviene del primer arco faríngeo, mientras que la musculatura de la lengua deriva de los somitos. La musculatura de la laringe también proviene de los arcos faríngeos. En cuanto a la musculatura de la columna vertebral, esta proviene del miotomo medial o interno del somito, en contraste con la musculatura de los miembros y la pared del cuerpo que se origina del miotomo externo. La musculatura cardíaca, por su parte, tiene su origen en el mesodermo lateral esplácnico.
El estudio del desarrollo de mioblastos en fetos humanos, de ratón y pollo, en regiones como la facial, lengua, columna vertebral, miembros, pared del cuerpo y musculatura cardíaca, se ha realizado mediante técnicas histológicas histoquímicas e inmunohistoquímicas para comprender mejor estos procesos.
Formación y Estructura de las Fibras Musculares Esqueléticas
Las células que forman el músculo esquelético se denominan fibras musculares o miofibras y son largas estructuras cilíndricas. Embriológicamente, el tejido muscular esquelético se desarrolla gracias a la fusión de varios mioblastos, o células musculares precursoras. Esta fusión en cadena de mioblastos da como resultado la característica célula muscular esquelética multinucleada, un fenómeno que también ocurre durante la vida fetal y postnatal. Debido a esto, se dice que estas células forman un sincitio estructural. Las fibras musculares pueden contener hasta varios miles de núcleos, la mayor parte de los cuales están localizados en la periferia, debajo de la membrana externa. En las células musculares esqueléticas, estos núcleos son ovalados y se ubican en la periferia de la célula.
Las fibras musculares tienen entre 10 y 100 µm de diámetro y pueden medir desde unos pocos milímetros hasta centímetros de longitud. Por ejemplo, en la cara anterior del muslo existen fibras musculares que pueden medir hasta un metro de longitud, mientras que las del músculo estapedio, en el oído medio, miden solo unos pocos milímetros de largo. El músculo sartorio, por su parte, puede tener fibras de 100 µm de diámetro y hasta 20 cm de longitud.
Están acompañadas por células satélite, ubicadas entre la lámina externa y el sarcolema. Las células satélite son células precursoras de células musculares y son las responsables de la capacidad regenerativa del tejido muscular esquelético.
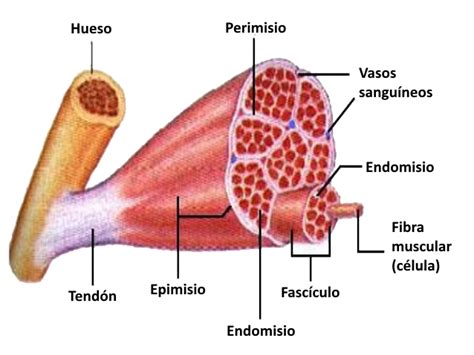
Organización del Tejido Conectivo
Cada fibra muscular está rodeada por una delgada capa de tejido conectivo llamada endomisio (membrana externa). Miles de estas fibras, envueltas por otra delgada capa de tejido conectivo llamada perimisio, forman un haz de fibras, al cual se le denomina fascículo o haz muscular. El perimisio es una capa ligeramente más gruesa que consta principalmente de colágeno tipo I y tipo III. Varios haces de grupos de fibras musculares se unen a un tendón en cada extremo y constituyen los músculos, que están rodeados por una membrana protectora llamada epimisio. Este tejido conectivo asociado une las fibras musculares en fascículos y también transporta fibras nerviosas y vasos sanguíneos (capilares) hacia las células musculares. Todo el músculo finalmente está rodeado por epimisio y se encuentra dentro de su fascia, un tejido conectivo compacto que separa el músculo de las estructuras circundantes.
Desarrollo de las Unidades Contráctiles y Membranas
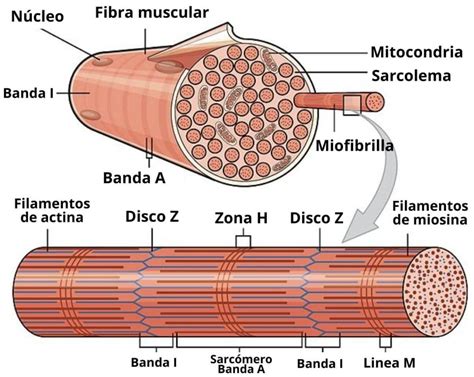
Miofibrillas y Sarcómero
Las fibras musculares se componen de miofibrillas, membranas y redes de citoesqueleto que anclan las fibrillas contráctiles al sarcolema. Las miofibrillas son haces de filamentos gruesos y delgados organizados en unidades funcionales repetitivas llamadas sarcómeros. La apariencia estriada de cada miofibrilla se debe a las diferencias en tamaño, densidad y distribución de los filamentos gruesos y delgados. Un sarcómero es la unidad funcional más pequeña de la fibra muscular, midiendo alrededor de 2.5 micrómetros de longitud en reposo, lo que significa que una miofibrilla puede contener aproximadamente 10,000 sarcómeros de un extremo a otro.
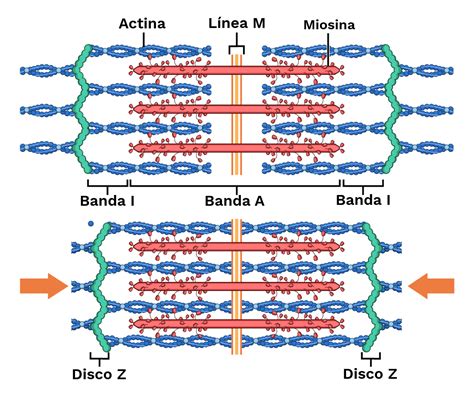
Cada sarcómero está formado por múltiples filamentos de proteínas llamadas actina y miosina, orientadas paralelamente entre sí. Los filamentos de actina y miosina se superponen en algunas partes generando varias bandas y zonas:
- El disco Z forma el límite de un sarcómero a cada extremo. Delgados filamentos de actina se proyectan en cualquier dirección desde un disco Z.
- La banda A está en el medio y corresponde a los filamentos de miosina que en sus extremos se superponen con los filamentos delgados de actina. Los polímeros de moléculas de miosina forman la banda A, oscura.
- La banda I se extiende desde la banda A de un sarcómero hasta la banda A del siguiente, conteniendo filamentos delgados, pero no filamentos gruesos. Las líneas Z marcan la frontera entre sarcómeros adyacentes.
- La región media del sarcómero carece de filamentos de actina y se denomina banda H, que bisecta la banda A.
- La línea M se encuentra ubicada en la mitad de la banda H y es perpendicular a los filamentos. En el centro de las bandas A corre la línea M donde se fijan los filamentos de miosina.
Los componentes primarios de las fibrillas de músculo esquelético son miosina y actina, así como tropomiosina y troponina asociadas con la actina.
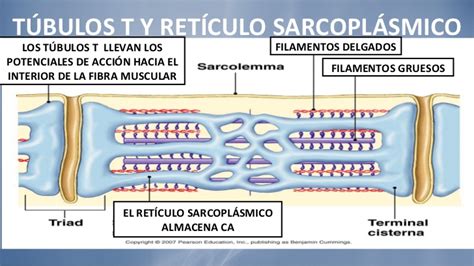
Retículo Sarcoplasmático (RS)
El retículo sarcoplasmático es un retículo endoplasmático liso modificado. Este complejo membranal se asemeja a una malla que rodea cada miofibrilla, formando una red tubular. A cada lado de un túbulo T, el retículo sarcoplasmático se ensancha y forma una cámara llamada cisterna terminal, que se une al túbulo T por medio de una estructura conocida como "pie". Un túbulo T y dos cisternas terminales forman una tríada.
Inicialmente, el retículo sarcoplasmático se desarrolla como un retículo endoplasmático, pero conforme el músculo se diferencia, se enriquece con proteínas específicas. Las cisternas terminales recogen el calcio que se usará en el ciclo de la contracción muscular. El RS controla el nivel de Ca2+ intracelular en el músculo esquelético, almacenándolo y liberándolo.
Tres proteínas purificadas inicialmente del retículo sarcoplasmático son la ATPasa de calcio (SERCA), calsecuestrina (CLQ) y el receptor a la rianodina (RyR). La SERCA es responsable de bombear calcio hacia la luz del RS durante la relajación, mientras que la CLQ es la proteína intraluminal quelante de calcio más prominente, y entre ambas aumentan la capacidad del RS para el calcio. La CLQ y RyR están acopladas funcionalmente en el RS de la unión.
Túbulos T (Transversales)
Los túbulos T (también llamados túbulos transversales) son invaginaciones a lo largo de la superficie de las células musculares esqueléticas. Sus paredes son continuas con las del sarcolema, lo cual significa que el túbulo T está expuesto a la matriz extracelular. Estos tubos estrechos se extienden en ángulos rectos a la superficie celular y forman vías llenas de líquido extracelular dentro de la fibra muscular. Como su membrana tiene las mismas propiedades generales que el sarcolema, los potenciales de acción son conducidos hasta llegar a la región de las tríadas, donde se inicia el proceso de acoplamiento excitación-contracción.
Los túbulos T forman una red continua que corre transversalmente por varias miofibrillas y las penetra a todos los niveles. Mantienen una composición de lípidos y proteínas diferente a la del sarcolema. Algunos estudios han proporcionado evidencias de que se originan como un compartimento tubular interno que después se funde con la membrana del sarcolema, mientras que otros sugieren que se forman por la fusión repetida de caveolas, ayudadas por la anfifisina. En el músculo esquelético diferenciado, la caveolina-3 se asocia con los túbulos T en desarrollo, aunque en el músculo maduro se restringe a las caveolas del sarcolema.
La caveolina-3, específica de músculo, puede ser requerida para generar la composición única de proteínas y lípidos del sistema tubular-T, y la anfifisina II es esencial para su organización y morfología normal. La anfifisina II se colocaliza con la anquirina y tiene un papel en anclar la ramificación de los túbulos y en organizar sus componentes proteicos.
Proteínas Accesorias y del Citoesqueleto en el Desarrollo Muscular
Los filamentos gruesos y delgados de las miofibrillas están acompañados de proteínas accesorias esenciales para la miogénesis y la función muscular. Estas proteínas mantienen la velocidad y el alineamiento de los filamentos durante el ciclo de la contracción muscular.
- Tropomodulina: Actúa como un casquete para la actina.
- Desmina: Es un filamento intermedio que forma una malla que rodea al sarcómero cerca de las líneas Z para unirlos entre sí y a la membrana plasmática.
- Nebulina: Es una proteína delgada y alargada que corre paralela a los filamentos delgados. Asiste a la α-actinina en la unión de los filamentos delgados a la línea Z y se cree que tiene importancia durante el desarrollo muscular. Probablemente, la longitud de la molécula de nebulina determina la longitud del hilo de actina-F conforme los filamentos delgados se desarrollan.
- Distrofina: Parece ligar los filamentos de actina a la lámina externa de la célula muscular. En el músculo esquelético, la distrofina del sarcolema se asocia a otras proteínas para formar un complejo que une el citoesqueleto de actina a la matriz extracelular. Esta unión es crucial para la función y supervivencia de la miofibrilla, ya que estabiliza el sarcolema durante los ciclos repetidos de contracción y relajación, y transmite la fuerza generada en los sarcómeros a la matriz extracelular. Las integrinas y el complejo de glicoproteínas asociada a la distrofina (DGC) ayudan a mantener este anclaje. Uno de sus componentes extracelulares, el β-distroglicano, proporciona un eslabón físico entre la matriz extracelular y el citoesqueleto intracelular.
Las proteínas estructurales del citoesqueleto que sostienen los filamentos intermedios, como las proteínas vimentina, desmina y nestina, forman un eslabón físico que en los músculos estriados conecta las subunidades contráctiles al sarcolema. Estos filamentos pueden tener un papel importante en la organización celular durante la miogénesis, así como en mantener la integridad estructural de las miofibras maduras, redistribuyendo el estrés producido por la actividad contráctil. En miofibras maduras, la expresión de vimentina está completamente regulada a la baja, mientras que la nestina, adyacente a la unión neuromuscular (NMJ) y la unión miotendinosa (MTJ), se expresa a bajos niveles.
El Acoplamiento Excitación-Contracción
Una fibra muscular esquelética puede ser muy larga, por lo que la señal que inicia la contracción debe distribuirse rápidamente a lo largo y hacia el interior de la célula para que todas sus regiones se contraigan simultáneamente. Para ello, la señal es propagada primero a lo largo del sarcolema y después conducida por los túbulos transversos o túbulos-T. Como la membrana del túbulo T tiene las mismas propiedades generales que el sarcolema, los potenciales de acción son conducidos hasta llegar a la región de las tríadas, donde se inicia el proceso que acopla la excitación con la contracción. Los túbulos-T están localizados entre la cisterna terminal adyacente al RS, formando una tríada compuesta por dos cisternas terminales y el túbulo-T.
Cuando el potencial de acción nervioso llega a la membrana muscular, esta sufre una despolarización que es transmitida a las profundidades de la célula por las membranas del sistema tubular-T. Cuando el potencial de acción de los túbulos es detectado por un receptor a la dihidropiridina (DHPR) localizado en la unión del túbulo con la cisterna del retículo sarcoplásmico, se produce la apertura de los receptores a la rianodina (RyR), que funcionan como canales para el calcio. Esta serie de eventos es llamada el 'acoplamiento excitación-contracción' (e-c). El DHPR en el túbulo-T detecta el voltaje a través de la membrana y su activación produce la liberación de Ca2+ del RS, mientras que el canal de liberación RyR/Ca2+ está localizado en el RS de la unión y es responsable de la liberación de Ca2+ de sus lugares de almacenamiento.
Para que ocurra la interacción entre la actina y la miosina que produce la contracción, debe haber calcio. Después de la contracción, el calcio debe ser eliminado, y la entrega y eliminación de este ion se lleva a cabo por el trabajo combinado del sistema tubular-T y el RS.
Tipos de Fibras Musculares Esqueléticas
Aunque no es directamente sobre desarrollo embrionario, la diferenciación de las fibras musculares esqueléticas es un resultado del proceso de desarrollo. Se pueden clasificar en:
- Fibras tipo I (oxidativas lentas): Especializadas en actividad aeróbica. Son pequeñas, contienen una elevada cantidad de mioglobina y se ven de color rojo en el tejido muscular fresco. Conforman unidades motoras de contracción lenta y resistentes a la fatiga.
- Fibras tipo IIa (oxidativas glucolíticas rápidas): Se ven de un color ligeramente más claro que las de tipo I en tejido muscular fresco. Contienen numerosas mitocondrias y poseen un contenido mayor de mioglobina que las de tipo IIb. Poseen gran cantidad de glucógeno, siendo capaces de realizar glucólisis anaeróbica. Forman unidades motoras de contracción rápida y resistentes a la fatiga. Son más resistentes a la fatiga que las de tipo IIb y se emplean en movimientos que requieren fuerza sostenida.
- Fibras tipo IIb (glucolíticas rápidas): Son fibras grandes que se ven de color rosa pálido en los tejidos frescos. Contienen menos mitocondrias y una menor cantidad de mioglobina. Aunque contienen un bajo nivel de enzimas oxidativas, exhiben una alta actividad de enzimas anaeróbicas y una gran cantidad de glucógeno. Son más propensas a la fatiga que las fibras de tipo I y IIa, formando unidades motoras de contracción y fatiga rápida.