La migración celular es un proceso biológico fundamental, indispensable tanto en el desarrollo embrionario como a lo largo de la vida del organismo. Desempeña un papel crucial en la formación, crecimiento y regeneración de los seres vivos. En organismos adultos, tiene gran importancia en procesos homeostáticos como la respuesta inmune y la reparación de tejidos. Sin embargo, la migración celular también puede contribuir a algunos procesos patológicos, incluyendo enfermedades vasculares, enfermedades inflamatorias crónicas y procesos metastáticos. Desde la fecundación, las células comienzan a multiplicarse, a diferenciarse y a madurar, debiéndose desplazar a sus regiones de destino mediante la migración para constituir lo que más adelante será un ser vivo.
Tipos de Migración Celular Durante el Desarrollo
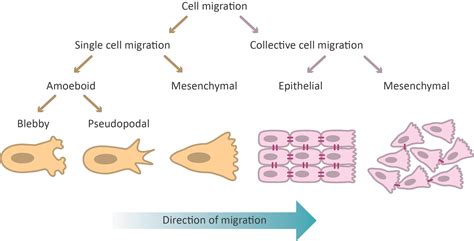
Durante el desarrollo, existen dos modos principales de movimiento celular: la migración individual y la migración colectiva.
Migración Individual
Las células que migran de forma individual son células procedentes de epitelios que, mediante una transición epitelio-mesénquima (EMT), experimentan cambios en la polaridad y adhesión celular. A través de este proceso, se delaminan y se convierten en células mesenquimales con capacidad de migrar de manera autónoma.
Migración Colectiva
La migración colectiva se entiende como el movimiento coordinado de varias células, ya sea formando parte de un grupo, de una fila o de una capa, que mantienen características similares a las células epiteliales. Este tipo de migración no se rige por una EMT como tal, ya que las células no pierden totalmente su identidad epitelial. Existen varios ejemplos de migración colectiva durante el desarrollo, como la gastrulación, y en procesos patológicos, como la metástasis.
Mecanismos Moleculares de la Migración Celular
En general, la migración celular se considera un proceso cíclico que requiere una compleja orquestación de eventos moleculares. Este ciclo comienza con la adquisición de una polaridad celular que genera una asimetría espacial entre la parte frontal y la trasera de la célula.
Polarización y Señalización Celular
Esta polarización se debe a la recepción de señales externas, tales como estímulos quimiotácticos, factores de crecimiento o proteínas de la matriz extracelular, las cuales determinan la dirección de la migración. Estas señales activan a receptores de membrana situados en el frente celular, como los receptores tirosina kinasas (RTK) EGFR y PDGF. Estos receptores se activan gracias a la fosforilación de su dominio intracelular por la unión de sus ligandos al dominio extracelular.
Otra proteína implicada en el establecimiento de la polaridad celular y en la adhesión y mantenimiento de epitelios es la E-cadherina. Esta proteína ejerce un papel importante en la migración colectiva durante el desarrollo: mientras que la disminución de sus niveles en el frente de células epiteliales promueve la migración, el aumento de su expresión la inhibe. Esto ocurre en procesos clave como el cierre del tubo neural o durante la gastrulación en el desarrollo embrionario.
5. Polaridad celular y basal
Formación de Proyecciones y Citoesqueleto
La polarización y la consecuente activación de las rutas de señalización de los receptores EGFR y PDGF implican la emisión de proyecciones transitorias en el frente celular. Estas prolongaciones o extensiones, conocidas como filopodios y lamelipodios, son responsables del movimiento celular.
Una molécula importante implicada en la formación de estas extensiones es la actina, un componente fundamental del citoesqueleto celular. La polimerización y despolimerización de la actina para la formación de proyecciones celulares depende de reorganizaciones del citoesqueleto. En este proceso están involucradas las proteínas pertenecientes a las Rho GTPasas, específicamente Rac1 y Cdc42. Varios estudios han demostrado que la activación de Rac1 o Cdc42 resulta en un incremento de la formación de lamelipodios y filopodios, respectivamente.
Adhesión y Retracción Celular
Una vez formados los lamelipodios y filopodios, deben anclarse al sustrato, ya sea a la matriz extracelular o a otras células adyacentes. Estos anclajes se denominan adhesiones focales o fibras de estrés. El proceso de unión de las proyecciones al sustrato se lleva a cabo por otro miembro de la familia de las Rho GTPasas, denominado Rho, a través de su interacción con actina.
Al mismo tiempo que se generan las adhesiones focales en las proyecciones del frente celular, la parte trasera de la célula debe retraerse para permitir el movimiento. En este proceso también se encuentra implicada la proteína Rho, la cual debe ser inhibida en la parte trasera para desestabilizar las uniones de la célula al sustrato, a la vez que ejerce su papel en la contracción del cuerpo celular mediante su interacción con la miosina.
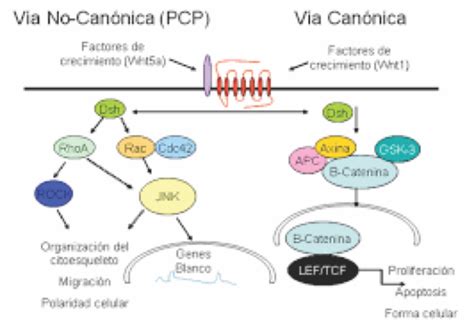
Regulación de las Rho GTPasas
Las Rho GTPasas deben estar bien reguladas para ejercer correctamente su papel de reorganización del citoesqueleto de actina durante la migración celular. Estas proteínas presentan dos estados conformacionales que dependen de su unión a GTP (activa) o a GDP (inactiva). La regulación de este ciclo de activación/inactivación depende de tres tipos de proteínas distintas:
- GEF (Guanine Nucleotide Exchange Factor): proteínas activadoras que catalizan la unión de las Rho GTPasas a GTP.
- GAP (GTPase Activating Proteins): proteínas inactivadoras implicadas en la hidrólisis del GTP y la posterior unión de las Rho GTPasas a GDP.
- GDI (Guanine Nucleotide Dissociation Inhibitor): proteínas inhibidoras que se mantienen unidas a las Rho GTPasas, previniendo su activación.
Los factores GEFs son los más estudiados en la activación de las GTPasas. Los miembros de la familia de las RhoGEF se caracterizan por poseer el complejo DH-PH, que interviene tanto en su localización en la membrana plasmática como en la regulación de su actividad catalítica. Un ejemplo destacado es la proteína Vav, que contiene un bucle de autoinhibición que regula negativamente su dominio catalítico DH, bloqueando la accesibilidad a las Rho GTPasas. Tras la estimulación de receptores tirosina kinasas, Vav es fosforilado, estimulando su dominio catalítico DH y permitiendo su interacción con las Rho GTPasas, como Rac1. En resumen, los receptores tirosina kinasas juegan un papel fundamental en la migración celular dirigida, ya que su activación por señales externas desencadena una cascada de señalización que, a través de GEFs y Rho GTPasas, controla la reorganización del citoesqueleto y, por tanto, la migración celular.
Ejemplos de Migración Celular en el Desarrollo Embrionario
Migración de las Células del Borde en Drosophila melanogaster
El estudio de la migración celular en modelos animales, como la mosca de la fruta Drosophila melanogaster, ha proporcionado valiosa información sobre los mecanismos moleculares en un organismo vivo. El sistema reproductivo de la hembra adulta de Drosophila está compuesto por dos ovarios, cada uno con varias ovariolas que contienen una hilera de cámaras huevo en distintos estadios del desarrollo. Las cámaras huevo están formadas por células nutricias y un oocito, rodeadas por células foliculares. En los extremos se localizan un par de células foliculares especializadas denominadas células polares.
En el estadio 8 del desarrollo, las células polares anteriores reclutan a células foliculares vecinas, formando el grupo de las células del borde (BCs). Estas BCs migran de forma colectiva a través de las células nutricias hasta contactar con el oocito en la parte posterior de la cámara huevo en el estadio 10. Esta migración se asemeja a una transición epitelio-mesénquima modificada, ya que las células foliculares pierden su polaridad epitelial, excepto las dos células polares que permanecen en el centro del grupo. Se establece una asimetría en el grupo de las BCs, con acumulación preferencial de F-actina en el frente de migración, donde se forman las extensiones de membrana necesarias para el movimiento.
La Rho GTPasa Rac1 está implicada en esta reorganización de actina, y su deficiencia resulta en defectos migratorios. Se ha observado una asimetría en la activación de Rac1, siendo mayor en el frente de las BCs, lo que es también afectado por la disminución de la expresión de PVR o EGFR. Esto sugiere que EGFR y PVR activan a Rac1 en el frente de las BCs, dirigiendo la migración hacia el oocito.
Estudios en este modelo han investigado el papel de proteínas como Coracle, homólogo de la proteína 4.1 de mamíferos, implicada en la reorganización del citoesqueleto de actina y la polaridad celular. Se observó que Coracle se localiza específicamente en las células polares anteriores durante la migración de las BCs, y deficiencias en su expresión alteran la distribución de la actina. Otros genes candidatos, como CG2887, CG33993 y vav, también han mostrado ser necesarios para la correcta migración de las BCs, con su función potencialmente ligada a la regulación del citoesqueleto de actina y, en el caso de CG2887 y CG33993, a la regulación de la proteína de polaridad celular Dlg.
La proteína Vav se localiza en las proyecciones de las BCs y su localización está regulada durante las fases de migración. Deficiencias en la expresión de Vav producen una distribución alterada de las proyecciones, aumentando su número y vida media en el frente. Estos hallazgos demuestran que Vav es esencial para la correcta distribución, estabilidad y longitud de las proyecciones frontales, lo que afecta directamente la velocidad y la trayectoria de la migración de las BCs. Deficiencias en Vav disminuyen la velocidad de migración y alteran la trayectoria, volviéndola no lineal, mientras que su sobreactivación puede resultar en una falta de migración.
Gastrulación y Formación de Capas Embrionarias en Humanos
Después de la implantación embrionaria, entre las semanas 4 y 5 de embarazo, el embrión en estado de blastocisto prolifera rápidamente y sufre cambios estructurales. El blastocisto, que es el estado del embrión a partir del quinto día de desarrollo, tiene una estructura de blástula con dos tipos celulares diferenciados: la masa celular interna (MCI) y el trofoectodermo.
La masa celular interna del blastocisto se convierte en una masa aplanada llamada disco embrionario, que se organiza en dos capas: el epiblasto y el hipoblasto. Este disco embrionario es el origen de todos los tejidos y órganos del futuro embrión. Las células del trofoectodermo se diferencian en estructuras que formarán la placenta.
El Proceso de Gastrulación
La gastrulación es el proceso mediante el cual el disco embrionario bilaminar pasa a ser un disco embrionario trilaminar, con tres capas embrionarias diferenciadas: ectodermo, mesodermo y endodermo. La gastrulación se inicia a partir de la blástula, cuando las células del epiblasto se dividen y proliferan rápidamente, necesitando migrar hacia nuevas localizaciones en el embrión. Esta migración celular lleva a las células del epiblasto a dirigirse hacia el hipoblasto y a desplazar a sus células, dando lugar a la tercera capa embrionaria (mesodermo) entre las dos anteriores.
El proceso de gastrulación humana es una etapa crucial del desarrollo embrionario, ya que a partir de las tres capas u hojas embrionarias se generarán todos los tejidos y órganos del cuerpo. En los embriones humanos, este proceso ocurre por invaginación, también llamada gastrulación por embolia, donde las células de la MCI del blastocisto se pliegan hacia el interior y migran, dando lugar a las capas embrionarias.
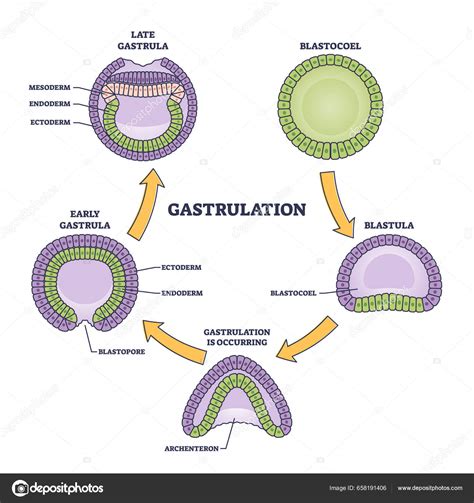
Las Capas Embrionarias y sus Derivados
Cada una de las tres capas embrionarias está destinada a formar un tipo de tejido diferente:
Ectodermo
Es la capa más externa que rodea al embrión y la primera en formarse a partir del epiblasto. Con el inicio de la organogénesis, las células del ectodermo se diferenciarán en:
- Ectodermo neural: formará el sistema nervioso (cerebro y médula espinal).
- Ectodermo superficial: formará los tejidos más superficiales, como la epidermis, el pelo, las uñas, las glándulas mamarias, las glándulas subcutáneas y el esmalte de los dientes.
- Un grupo de células ectodérmicas, los amniocitos, formarán parte del saco amniótico.
Mesodermo
Es la capa intermedia, que se forma a partir de la migración de las células del epiblasto. El mesodermo es la hoja embrionaria que formará la mayoría de los tejidos y órganos del futuro feto. Se diferencia en:
- Notocorda: se sitúa en el eje longitudinal del embrión y actúa como sostén, siendo fundamental para la formación del tubo neural.
- Mesodermo paraxial: se desarrolla en el dorso del embrión, formando los pares de somitas que originarán el tejido muscular, esquelético, cartilaginoso y la dermis.
- Mesodermo intermedio (nefrotomo): dará lugar a los riñones y otros componentes del sistema urogenital.
- Mesodermo lateral: originará el sistema sanguíneo y cardiovascular, así como el endotelio vascular y las membranas del mesotelio.
Endodermo
Es la capa más interna. Con la diferenciación del cuerpo del embrión, se divide en dos partes:
- Endodermo embrionario: dará lugar a los órganos internos del cuerpo, es decir, aquellos que forman el aparato digestivo y el sistema respiratorio (faringe, estómago, intestino, hígado, páncreas, vesícula biliar, bronquios, vejiga urinaria, etc.).
- Endodermo extraembrionario: la parte que queda fuera del embrión y que forma el saco vitelino, una estructura encargada de nutrir y aportar oxígeno al embrión durante las primeras semanas de desarrollo.
Estas dos partes endodérmicas quedan comunicadas por un orificio ancho que se convertirá en el cordón umbilical.
Importancia y Riesgos de la Gastrulación
La gastrulación es un proceso que ocurre en una etapa muy temprana del desarrollo embrionario, alrededor de la quinta semana de embarazo. Cualquier alteración o anomalía durante la formación de las tres capas embrionarias podría conducir a la detención del desarrollo del embrión o a la aparición de defectos congénitos graves, subrayando su importancia crítica.
Neurulación
La neurulación es el proceso embriológico en el que se forma el tubo neural del embrión, la estructura precursora del sistema nervioso central, que dará lugar al cerebro y la médula espinal. El tubo neural se origina a partir de las células del ectodermo que forman la placa neural. Sus células se pliegan y se fusionan formando una estructura cilíndrica. El proceso de neurulación ocurre después de la gastrulación y suele corresponder con la quinta semana de embarazo.
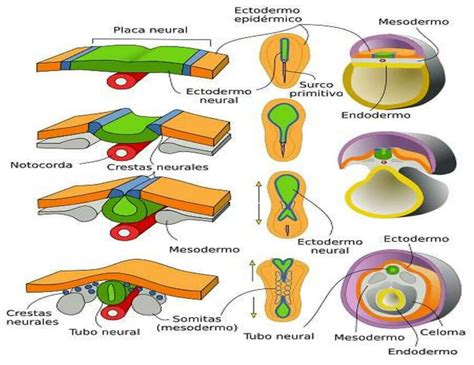
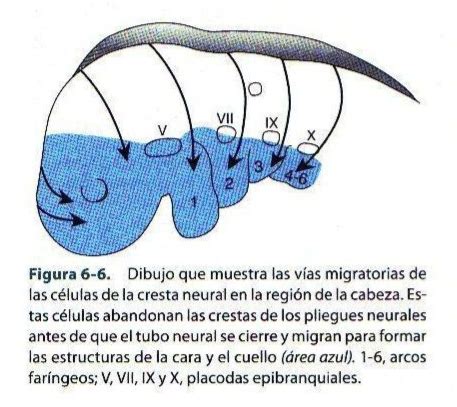
Migración de las Células de la Cresta Neural
Las células de la cresta neural son una estructura embrionaria de los vertebrados formada por células migratorias que generan una sorprendente diversidad de tejidos, incluyendo células del sistema nervioso periférico, del sistema cardiovascular, células pigmentarias de la piel y gran parte del tejido conectivo de la cabeza, los huesos y los cartílagos.
Durante su desarrollo, las células de la cresta neural muestran un comportamiento migratorio peculiar, similar a un juego de "pilla-pilla" o "persecución y huida". Estas células "persiguen" a otros tipos de células, las de las placodas (láminas que forman los órganos sensoriales), y estas últimas huyen cuando están a punto de ser alcanzadas. La "persecución" se produce en presencia de una proteína de señalización, la Sdf1, mientras que la "huida" la provoca otra proteína, la N-cadherina.
La interacción física entre estas células, incluyendo fuerzas de cizallamiento, empujes y tirones, coordina la migración de las láminas de células. Estudios sobre la migración de las células mesenquimatosas de la cresta neural (NCC) en modelos como Xenopus laevis han utilizado microscopia de secuencia temporal y microscopia de fuerzas de tracción para observar y medir las fuerzas ejercidas por las células. Al encontrarse con otras células, ocurre un fenómeno de inhibición de las protrusiones de las células mesenquimatosas y el alejamiento de las células, denominado inhibición del movimiento por contacto (CIL). Aunque la CIL es necesaria en la migración colectiva mesenquimatosa, no explica la totalidad del fenómeno, lo que resalta la complejidad de estos procesos.
El Papel de las Proteínas YAP y las Fuerzas Mecánicas
Investigaciones recientes han destacado el papel de las proteínas YAP, activadores transcripcionales implicados en el mantenimiento de tejidos adultos y la embriogénesis, como fundamentales para la correcta formación del embrión in vivo. Durante la gastrulación, estas proteínas son cruciales para que las células migren a sus destinos finales y para que el eje del embrión se forme correctamente. Yap aparece en el epicentro del control de mecanismos morfogenéticos complejos, con un sistema de regulación ligado a la morfología de la célula y su geometría nuclear.
Durante la formación de un organismo, los tejidos en desarrollo están recubiertos por una matriz extracelular especializada llamada membrana basal, que es capaz de alterar las fuerzas que actúan en su interior, regulando la morfogénesis (el proceso por el que el embrión adquiere su forma) y la homeostasis de los tejidos. Identificar las señales responsables de la migración celular, especialmente aquellas que dependen de fuerzas mecánicas, es importante no solo para comprender las bases de la formación de embriones y órganos, sino también para identificar dianas terapéuticas contra procesos de migración incontrolada, como la metástasis cancerígena y otras enfermedades relacionadas con problemas durante el desarrollo embrionario.
Incluso después de la formación del individuo adulto, los tejidos pueden sufrir daños. Por ello, se estudian los factores mecánicos durante la migración celular en el desarrollo cerebral y en la regeneración neural como herramientas fundamentales para entender estos procesos. Por ejemplo, las células de Cajal-Retzius (CRcs) en el cerebro embrionario de ratón muestran tasas de migración diferenciales influenciadas por factores físicos, evidenciando la interacción entre la mecánica y la biología celular en el desarrollo.
tags: #migracion #celular #embrionaria