La hemoglobina (Hb) es una molécula de gran relevancia científica, cuya importancia en la biología ha propiciado sorprendentes descubrimientos a lo largo de los años. Desde el siglo XIX, cuando se realizaron los primeros estudios científicos, hasta la actualidad, continúan apareciendo hallazgos notables, como las nuevas globinas neuroglobina y citoglobina, y sus llamativas interacciones con el óxido nítrico. El estudio de las hemoglobinopatías, por su parte, representa un gran desafío para la medicina moderna, al buscar aplicar los resultados de la investigación científica básica en el servicio a los pacientes.
Un ejemplo sobresaliente de la relevancia del proceso evolutivo y la eficiencia de los sistemas biológicos se manifiesta en los eritrocitos. Una de sus funciones vitales es la participación en el intercambio gaseoso de oxígeno (O2) y dióxido de carbono (CO2) entre los pulmones y los tejidos. La hemoglobina es el componente fundamental de este proceso.
La Hemoglobina: Un Modelo Molecular Crucial en la Biología
Las hemoglobinas son proteínas globulares presentes en los hematíes en altas concentraciones. Su función principal es fijar oxígeno en los pulmones y transportarlo por la sangre hacia los tejidos y células que rodean el lecho capilar del sistema vascular. Al volver a los pulmones, desde la red de capilares, la hemoglobina actúa como transportador de CO2 y de protones.
A lo largo de la historia, la hemoglobina ha desempeñado un papel fundamental en la química, la biología y la medicina. En 1849, se convirtió en la primera proteína en ser cristalizada y asociada con una función fisiológica específica. La diferencia morfológica entre los cristales de hemoglobina de diferentes organismos proporcionó por primera vez evidencia contundente sobre la especificidad en la expresión proteica entre las especies. Además, fue una de las primeras proteínas cuyo peso molecular se determinó correctamente. En 1958, se logró sintetizarla in vitro como la primera proteína eucariota, lo que permitió comprobar que el mecanismo de síntesis proteica en eucariotas es similar al de Escherichia coli. Su estructura se estableció en 1960. El ARN mensajero de la globina fue el primer mensajero eucariota en ser aislado y en tener una secuencia nucleotídica determinada.
El descubrimiento de que la anemia de células falciformes es causada por el reemplazo de uno solo de los 287 residuos de aminoácidos, presentó por primera vez indicios de que una mutación puntual en un gen estructural puede causar la sustitución de un aminoácido en la proteína codificada por este gen y, por ende, provocar una enfermedad. El estudio de la hemoglobina también impulsó el desarrollo de nuevos y sofisticados métodos físicos, así como el establecimiento de importantes teorías sobre cooperatividad y alosterismo. De igual forma, la transición de la síntesis de hemoglobina desde la vida fetal a la adulta es un claro ejemplo de diferenciación celular. El estudio de las hemoglobinas anormales ha permitido conocer la estrecha relación entre los errores genéticos, los defectos proteicos y las manifestaciones clínicas. Finalmente, la distribución de ciertas hemoglobinas anormales como la HbS en regiones endémicas de malaria ilustra los mecanismos naturales de la evolución y adaptación antropológica (polimorfismo balanceado).
Genética y Biosíntesis de la Hemoglobina
La biosíntesis de la hemoglobina está estrechamente relacionada con la eritropoyesis. La expresión genética y el contenido de hemoglobina acompañan la diferenciación de las unidades formadoras de colonias eritroides (UFC-E) en precursores eritroides. Cada una de las cadenas polipeptídicas de la Hb cuenta con genes propios: α, β, δ, γ, ε.
Organización de los Genes de Globina
- El grupo α-globina (que incluye los genes de la cadena ζ) se localiza en el brazo corto del cromosoma 16.
- El grupo β-globina (que incluye los genes de las cadenas Gγ, Aγ, δ y ε) se localiza en el brazo corto del cromosoma 11.
Todos los genes funcionales de la globina comparten una estructura general que consiste en tres exones (secuencias codificadoras) y dos intrones o sectores interpuestos (secuencias que no se traducen). La región promotora, que precede al punto de comienzo de la transcripción, incluye aproximadamente 100 pares de bases. Tres secuencias de esta región se fijan a la ARN polimerasa, la enzima que cataliza la síntesis de ARN mensajero. Dos secuencias clave en la iniciación de la transcripción son TATA y CAT; las mutaciones que las afectan limitan la transcripción de ARNm. La porción distal del tercer exón (AATAAA) finaliza la transcripción. Solamente entre el 5% y el 10% del material genético de los eritroblastos se transcribe, y los genes de la globina pertenecen a esta fracción.
Síntesis de ARN y Traducción
La síntesis de ARN se lleva a cabo bajo la influencia de grupos enzimáticos denominados ARN polimerasas. La transcripción primaria del ARNm incluye copias de toda la secuencia del ADN genómico (intrones y exones). Antes de su transporte al citoplasma, el pre-ARNm se procesa mediante el clivaje del extremo 5', la separación de las secuencias transcritas de los intrones y la poliadenilación del extremo 3'. Este último paso es esencial para el transporte y la estabilización citoplasmática del ARNm. La separación implica la formación de asas en el pre-ARNm, de manera que los extremos distales de los exones (puntos dadores) se acerquen a los proximales de los subsiguientes exones (puntos receptores). Posteriormente, los intrones sufren clivaje enzimático y los puntos dadores y receptores se sellan. Los puntos de consenso son secuencias de nucleótidos adyacentes que perfeccionan la síntesis del ARNm. Las mutaciones que involucran tanto los puntos de unión como los de consenso alteran la separación y crean ARNm anormales.
La causa más común de las hemoglobinopatías es la mutación puntual, es decir, la sustitución de un nucleótido de ADN por otro, lo que modifica el código genético y puede inducir un cambio en un aminoácido de la globina resultante. Por ejemplo, en la anemia de células falciformes (HbS), el ácido glutámico se reemplaza por una valina en el aminoácido 6, que ocupa el tercer lugar del helicoide A de la cadena β: β6 (A3) Glu → Val.
La traducción es un proceso ribosómico en el que se sintetiza una cadena polipeptídica de acuerdo con un patrón proporcionado por la secuencia de codones del ARNm. Incluye cuatro etapas:
- Activación: formación del ARN de transferencia (ARNt).
- Iniciación: el ARNt que contiene metionina se alinea con el codón iniciador AUG en el ARNm del ribosoma.
- Elongación: cada anticodón del ARNt se adosa a cada codón del ARNm. Los aminoácidos del ARNt se adosan mediante un puente peptídico a otro aminoácido ya unido al ribosoma.
- Terminación: se produce cuando se llega a un codón de finalización UAA, la cadena polipeptídica se completa y se separa del ribosoma.
Los polipéptidos libres forman de inmediato dímeros αβ y tetrámeros α2β2.
Regulación de la Eritropoyesis y Factores Asociados
La maduración de proeritroblastos a eritrocitos está controlada positivamente por la hormona polipeptídica eritropoyetina, que promueve tanto la proliferación como la supervivencia de los precursores eritroides. En la superficie de estos precursores, se encuentran receptores que promueven la apoptosis y que son estimulados por enzimas denominadas caspasas. La activación de estos receptores (o la privación de eritropoyetina) conduce al clivaje, por parte de las caspasas, de una proteína regulatoria nuclear llamada GATA-1, indispensable para el proceso eritropoyético, lo que a su vez conduce a apoptosis y detención de la maduración. Estos receptores de «muerte celular» se han denominado sistema Fas/FasL.
La investigación continua sobre los genes sobre los cuales actúa el factor de transcripción genético GATA-1 condujo al descubrimiento de un gen expresado en altos niveles en el eritroblasto, que codifica una proteína chaperona denominada AHSP (proteína estabilizante de cadena alfa). Esta proteína se une específicamente a las cadenas α, y su papel está relacionado con la estabilización de dichas cadenas, evitando que se precipiten formando inclusiones citoplasmáticas (cuerpos de Heinz) que afectan a la membrana celular y conducen a la lisis eritrocitaria.
Síntesis y Degradación del Grupo Hemo
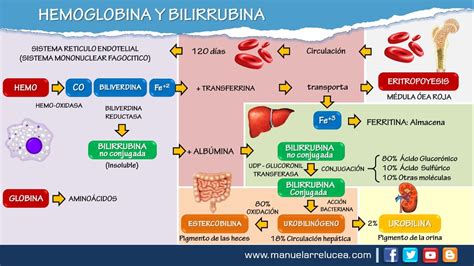
El grupo hemo se sintetiza en casi todos los tejidos, pero su síntesis es más pronunciada en la médula ósea y el hígado, debido a la necesidad de incorporarlo en la Hb y los citocromos, respectivamente. Es una molécula plana que consta de un hierro ferroso y un anillo tetrapirrólico, la protoporfirina IX. El hemo es un factor fundamental en la regulación de la tasa de síntesis de la globina. Su principal efecto se ejerce en la iniciación de la traducción, donde bloquea la acción de un inhibidor de la producción de globina. También participa en la transcripción y el procesamiento del ARNm. Su papel en la síntesis proteica en mamíferos se extiende más allá del eritrocito; en el tejido hepático y cerebral se han demostrado sustancias que dependen del hemo para comenzar la producción de proteínas.
Normalmente, los eritrocitos envejecidos se degradan hacia el día 120 de vida en la médula ósea, el hígado y el bazo. Sin embargo, en algunas circunstancias, los eritrocitos sufren lisis intravascular, liberando hemoglobina que puede ser tóxica para los tejidos a menos que se remueva rápidamente. La haptoglobina (Hp) es una proteína plasmática que se une a la Hb libre, formando un complejo Hp-Hb. Este complejo es reconocido por una proteína situada en la superficie de los macrófagos y monocitos denominada CD163, lo que permite su digestión y la subsecuente liberación de hierro y bilirrubina. La expresión de Hp y CD163 está regulada por proteínas de fase aguda como la interleucina 6 (IL-6), sugiriendo que las enfermedades inflamatorias crónicas se relacionan con alteraciones del metabolismo del hierro.
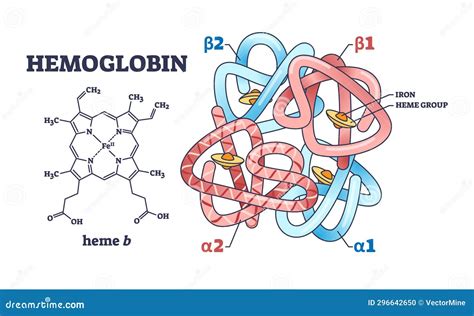
Estructura de la Hemoglobina y sus Cadenas Globínicas
La hemoglobina es una proteína compleja cuyas cuatro cadenas polipeptídicas contienen cada una un grupo prostético hemo. Un grupo prostético es la porción no polipeptídica de una proteína. El hemo es una molécula de porfirina que contiene un átomo de hierro en su centro, específicamente una protoporfirina IX, con dos grupos ácidos propiónicos, dos vinilos y cuatro metilos como cadenas laterales unidas a los anillos pirrólicos.
El átomo de hierro se encuentra en estado de oxidación ferroso (+2) y puede formar cinco o seis enlaces de coordinación dependiendo de la unión del O2 (u otro ligando) a la Hb (oxiHb, desoxiHb). Cuatro de estos enlaces se producen con los nitrógenos pirrólicos de la porfirina en un plano horizontal. El quinto enlace de coordinación se realiza con el nitrógeno del imidazol de una histidina denominada histidina proximal. Finalmente, el sexto enlace del átomo ferroso es con el O2, que además está unido a un segundo imidazol de una histidina denominada histidina distal. Tanto el quinto como el sexto enlace se encuentran en un plano perpendicular al plano del anillo de porfirina.
Las cadenas polipeptídicas α contienen 141 aminoácidos, mientras que las no α (β, γ, δ) contienen 146 y difieren en la secuencia de aminoácidos. La estructura primaria de las cuatro cadenas de Hb normales se conoce desde hace décadas. La estructura secundaria es muy similar: cada una exhibe 8 segmentos helicoidales designados con las letras A a la H. Entre ellos se encuentran 7 segmentos no helicoidales: NA, AB, CD, EF, FG, GH y HC. Esta distinción es fundamental, ya que los segmentos helicoidales son rígidos y lineales, mientras que los no helicoidales son flexibles. Dado que el hierro del hemo forma un puente covalente con la histidina proximal (F8) y el O2 se une de forma covalente al hemo y a la histidina distal (E7), el hemo queda suspendido en una hendidura no polar entre los helicoides E y F.
La difracción de rayos X de alta resolución permitió conocer la naturaleza de los contactos intercatenarios de la Hb. Los contactos entre cadenas semejantes (α1α2 y β1β2) son limitados y de escasa importancia. Los principales contactos son α1β1 y α1β2, los cuales determinan dos estructuras cuaternarias: una para la oxiHb y otra para la desoxiHb. La parte porfirínica del hemo se sitúa dentro de una bolsa hidrofóbica que se forma en cada una de las cadenas polipeptídicas. Las estructuras obtenidas por difracción de rayos X muestran que en la bolsa del hemo existen unas 80 interacciones entre 18 aminoácidos y el hemo. La mayoría de estas interacciones no covalentes se presentan entre cadenas apolares de aminoácidos y las regiones no polares de la porfirina.
La Familia Génica de las Globinas y su Evolución
La globina es una familia de proteínas estructurales esenciales que forman parte de la hemoglobina, una molécula responsable del transporte de oxígeno en la sangre de vertebrados. Las globinas son proteínas de naturaleza globular que contienen regiones capaces de unirse al grupo hemo, el cual posee un átomo de hierro. Las cadenas globínicas son subunidades polipeptídicas específicas que conforman la hemoglobina. Estas cadenas se organizan en dímeros y tetrámeros, dependiendo del tipo de globina y de la etapa del desarrollo en la que se encuentren. Las globinas tienen una función crítica en el transporte y almacenamiento de oxígeno, permitiendo a los organismos aerobios mantener sus funciones metabólicas.
Los genes se duplican en tiempos evolutivos, y esta duplicación puede conducir a la formación de familias de genes relacionados. Una familia de genes es un conjunto de varios genes similares, formados por la duplicación de un solo gen original, que generalmente tienen funciones bioquímicas similares. Una de estas familias son los genes de las subunidades de hemoglobina humana. Los 10 genes se encuentran en dos grupos en diferentes cromosomas, llamados loci α-globina y β-globina. Los genes se categorizan en familias basándose en secuencias de nucleótidos o proteínas compartidas. Las técnicas filogenéticas pueden ser utilizadas como una prueba más rigurosa. Las posiciones de los exones dentro de la secuencia codificante pueden usarse para inferir ascendencia común. Conocer la secuencia de la proteína codificada por un gen puede permitir a los investigadores aplicar métodos que encuentran similitudes entre secuencias proteicas, proporcionando más información que las similitudes o diferencias entre secuencias de ADN. Si los genes de una familia de genes codifican proteínas, el término familia de proteínas se usa a menudo de manera análoga a la familia de genes.
La expansión o contracción de familias de genes a lo largo de un linaje específico puede deberse al azar o ser el resultado de la selección natural, una distinción a menudo difícil de establecer en la práctica. Por el contrario, los complejos génicos son simplemente grupos de genes estrechamente unidos, a menudo creados a través de la duplicación génica (a veces llamada duplicación segmentaria si los duplicados permanecen uno al lado del otro).
Globinas Embrionarias, Fetales y Adultas: Un Desarrollo Programado
Las cadenas globínicas son las subunidades polipeptídicas que componen las moléculas de hemoglobina. La información genética para la síntesis de las diversas cadenas de globinas está contenida en diversos cromosomas. Durante el desarrollo embrionario, se sintetizan específicamente las globinas zeta (ζ) y épsilon (ε). Dos cadenas γ-globina se combinan con dos cadenas ζ-globina para formar la hemoglobina Portland embrionaria. En la etapa fetal predominan las cadenas alfa (α) y gamma (γ). Las alteraciones en las cadenas globínicas pueden tener consecuencias clínicas importantes y dar lugar a enfermedades hematológicas, conocidas como hemoglobinopatías.
Las talasemias son trastornos hereditarios caracterizados por la producción deficiente de cadenas globínicas. La drepanocitosis (anemia de células falciformes o drepanocítica) es una hemoglobinopatía causada por una mutación en la cadena beta de la globina. La evaluación de las cadenas globínicas es fundamental en el diagnóstico de hemoglobinopatías. Las diferencias entre haplotipos en portadores de hemoglobina S (HbS) radican en la cantidad de hemoglobina F que poseen; a mayor porcentaje de hemoglobina F y menor hemoglobina S, mejor pronóstico.
ADN, Cromosomas y Genes
Otras Globinas: Mioglobina, Neuroglobina y Citoglobina
Hasta hace poco tiempo se creía que la hemoglobina y la mioglobina eran las únicas sustancias que contenían globina en los seres superiores, pero hoy día se conocen dos más que poseen globina en su constitución: la citoglobina y la neuroglobina.
Mioglobina
La mioglobina presenta una estructura menos compleja que la hemoglobina. Está constituida por un grupo proteico globular con una sola cadena peptídica de 153 aminoácidos. Su disposición espacial es secundaria y presenta 8 hélices alfa. Esta estructura proteica ubica de manera estratégica los aminoácidos hidrofóbicos hacia el interior, mientras que los aminoácidos hidrofílicos o polares se encuentran hacia el exterior, un diseño perfecto para albergar el grupo hemo en su interior (parte hidrofóbica). La mioglobina tiene una mayor afinidad por el oxígeno que la hemoglobina, liberando O2 solamente a muy bajas tensiones tisulares.
Citoglobina
Descubierta en 2001, la citoglobina es un tipo de hemoglobina que se diferencia por ser hexacoordinada, a diferencia de la hemoglobina y la mioglobina que son pentacoordinadas. Está presente en la mayoría de los tejidos, especialmente en el tejido conectivo, y se ha encontrado en la retina. Se cree que esta proteína puede influir en la protección contra estados de hipoxia y estrés oxidativo en los tejidos.
Neuroglobina
Su descubrimiento se realizó en 2000. La neuroglobina es un monómero que posee 150 aminoácidos, siendo muy similar a la mioglobina. Tiene preferencia por el tejido nervioso, de allí su nombre. Se piensa que la neuroglobina tiene capacidad para fijar oxígeno, monóxido de carbono y óxido nítrico. Aunque su función aún no se sabe con certeza, se cree que está relacionada con la regulación de la hipoxia e isquemia cerebral, actuando como un neuroprotector. Debido a su estructura similar a la hemoglobina y la mioglobina, se especula que podría participar en el suministro de oxígeno a nivel neuronal. También se cree que puede eliminar los radicales libres y el nitrógeno que se producen en la cadena respiratoria.
tags: #gamilia #genica #globina #embrionaria