Las células madre embrionarias (ESC, por sus siglas en inglés) han capturado la imaginación tanto de científicos como de no científicos. Estas células son cruciales para entender el desarrollo de un organismo y tienen un vasto potencial en la investigación médica. Pero, ¿qué son exactamente, cómo se obtienen y por qué los investigadores en todo el mundo trabajan tan diligentemente para comprenderlas mejor?
Fundamentos de la Diferenciación Celular y la Potencia
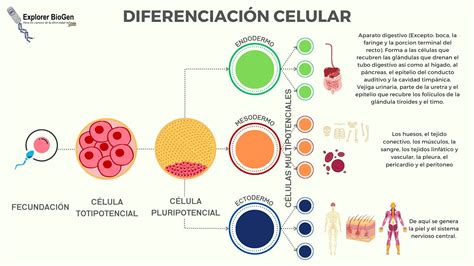
La diferenciación celular es el proceso por el cual las células adquieren una forma y una función determinada durante el desarrollo embrionario o la vida de un organismo pluricelular, especializándose en un tipo celular específico. En la inmensa mayoría de los organismos pluricelulares, no todas las células son idénticas; por ejemplo, las células que forman la piel en el ser humano son diferentes de las que componen los órganos internos. Sin embargo, todos los diferentes tipos celulares derivan de una sola célula inicial o cigoto, procedente de la fecundación de un ovocito II por un espermatozoide, gracias a la diferenciación celular.
La diferenciación es un mecanismo mediante el cual una célula no especializada se especializa en numerosos tipos celulares que forman el cuerpo, como los miocitos (células musculares), los hepatocitos (células del hígado) o incluso las neuronas (células del sistema nervioso). Durante la diferenciación, ciertos genes son expresados mientras que otros son reprimidos. Este proceso es intrínsecamente regulado gracias al material epigenético de las células, afectando cambios en numerosos aspectos de la fisiología celular como el tamaño, la forma, la polaridad, la actividad metabólica, la sensibilidad a ciertas señales y la expresión de genes.
Clasificación de las Células en Mamíferos
Las células de los mamíferos se pueden clasificar en tres categorías:
- Las células de la línea germinal: destinadas a la formación de gametos (óvulos o espermatozoides), son las únicas capaces de transmitir su material genético a las generaciones siguientes.
- Las células somáticas: la mayoría de las células, son diploides (poseen dos copias de cada cromosoma) y poseen su propia copia del genoma, excepto ciertas células que han perdido su núcleo celular durante su diferenciación, como los hematíes.
- Las células madre.
Potencia Celular
Cualquier célula que presente un nivel de potencia celular o capacidad de diferenciación, es lo que se denomina célula madre. Estas pueden clasificarse según su capacidad de diferenciación:
- Una célula capaz de producir la totalidad de tipos celulares, incluyendo los tejidos extra-embrionarios o placentarios, se le denomina una célula totipotente.
- Si la célula solo puede diferenciarse en linajes celulares presentes en el adulto, se denomina una célula pluripotente. Estas células pueden diferenciarse en cualquier tipo de célula del cuerpo.
- Una célula multipotente solo puede diferenciarse en pocos tipos celulares, generalmente del mismo linaje celular.
- Finalmente, cuando la célula solo puede producir un único tipo de célula, se le denomina unipotente.
Las células madre son capaces de cambiar y transformarse en otros tipos de células que se encuentran en el cuerpo; este proceso es la diferenciación celular y está a cargo del desarrollo de todas las células del cuerpo. En las células en desarrollo, existen diferentes niveles de potencia celular, es decir, la habilidad de la célula de diferenciarse en otros tipos celulares. Una mayor potencia implica una mayor cantidad de linajes celulares que puede producir.
El Origen de las Células Madre Embrionarias
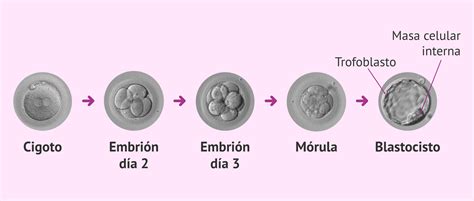
El desarrollo comienza cuando un espermatozoide fecunda un óvulo y crea una sola célula que puede potencialmente formar un organismo entero. Durante la primera hora después de la fecundación, esta célula huevo o cigoto se divide en varias células idénticas. En el ser humano, alrededor de cuatro horas después de la fecundación y después de varios ciclos celulares, estas células comienzan a especializarse y formar una esfera que crece llamada blastocisto.
El blastocisto es una estructura embrionaria presente en las etapas tempranas del desarrollo de mamíferos, antes de la implantación en el útero materno. Está compuesto por entre 64 y 100 células que rodean una cavidad central. Antes de que el embrión alcance este estadio, todas las células que lo forman son equivalentes y totipotentes, es decir, son capaces de formar todas las estructuras del embrión. Pero con la formación del blastocisto ocurre la primera distinción entre células.
Esta esfera posee una capa de células externas (las células periféricas) y un grupo de células internas, llamadas células de la masa interna, que formarán todos los tejidos del cuerpo humano. Como consecuencia de esta primera decisión de linaje que ocurre en el blastocisto, se establecen dos poblaciones de células: las que darán lugar al trofectodermo (origen de la futura placenta) y las que formarán la masa celular interna, origen del resto del embrión y del organismo adulto. Las células de la masa celular interna son pluripotentes: pueden diferenciarse en cualquier tipo de célula en el cuerpo. A pesar de esto, estas células solas no pueden formar individualmente un organismo entero, por lo que son calificadas como pluripotentes.

La segregación de estas células es clave ya que, por un lado, las células embrionarias pierden su totipotencialidad y, por otro, se genera un tejido, el trofectodermo, que es característico de mamíferos. De este tejido se origina el trofoblasto que, tras diferenciarse en varios tipos celulares, forma la placenta y las membranas amnióticas. Es ahí donde juega un papel fundamental el gen Cdx2, esencial en la segregación del trofectodermo frente a la masa celular interna. Los científicos han identificado un elemento regulador de la función de Cdx2 que dirige la expresión de un gen marcador de manera restringida al trofectodermo, siendo esencial para comprender la información que recibe y procesa Cdx2 para activarse y ejercer su función.
Cultivo de Células Madre Embrionarias en Laboratorio
Cuando se obtiene la masa celular interna de un blastocisto de ratón y se le dan los nutrientes adecuados, es posible cultivarla en el laboratorio. El proceso de maduración y especialización celular, que normalmente se llevaría a cabo en el embrión, se detiene. En cambio, las células se multiplican para producir más células desdiferenciadas parecidas a la masa celular interna. Estas células que se cultivan en el laboratorio se llaman células madre embrionarias (ES). Las células madre embrionarias pueden multiplicarse y son capaces de crear otros tipos de células más especializadas.
Investigación y Aplicaciones de las Células Madre Embrionarias
Las células madre embrionarias humanas (ESC) son células pluripotentes, lo que significa que pueden fabricar cualquier otra célula del cuerpo. Se utilizan extensamente en la investigación para entender cómo los blastocistos crecen hasta convertirse en células adultas y qué señales dirigen a las células madre a diferenciarse en células especializadas.
Modelos Quiméricos y Ratones Genéticamente Modificados
Las células madre de ratón pueden regresarse a un blastocisto de ratón y este blastocisto puede regresarse al útero de un ratón hembra para su crecimiento. Las células inyectadas toman parte en el desarrollo del embrión y el cachorro nace con una mezcla de células, (a) del blastocisto huésped y (b) de las células que provienen de las células ES inyectadas. A este nuevo ratón con células de dos orígenes diferentes se le llama una quimera. Se ha aprendido mucho sobre el desarrollo embrionario y la enfermedad creando ratones quimera con ESC genéticamente modificadas. Estos ratones permiten a los investigadores analizar cómo contribuyen genes específicos a las funciones y enfermedades celulares.
Las quimeras pueden heredar los genes de las células madre embrionarias a su descendencia. Los investigadores pueden entonces alterar los genes de las células ES en el laboratorio, regresar las células a los blastocistos y producir nuevos ratones con genes alterados o “mutaciones”. Los científicos utilizan ratones alterados genéticamente hechos a partir de células ES para estudiar genes involucrados en muchas enfermedades humanas. Por ejemplo, se han hecho ratones con mutaciones encontradas en cánceres humanos. Estos ratones pueden estudiarse para entender más sobre cómo crecen los cánceres y para probar posibles fármacos. El gran valor de esta técnica para la ciencia y la medicina fue reconocido en el año 2007 cuando se otorgó el Premio Nobel de Medicina a tres científicos pioneros en el uso de células ES para crear ratones genéticamente modificados.
Uso de Células Madre Embrionarias Humanas
Los científicos también han hecho células ES a partir de la masa celular interna de embriones humanos, utilizando la misma técnica que se utiliza en ratones para aislar células ES. Aunque no está permitido inyectar células ES humanas de regreso en los blastocistos por razones éticas y morales, estas células pueden diferenciarse en el laboratorio para formar muchos tipos de células especializadas. Al utilizar células ES humanas para producir células especializadas como las neuronas o las células de corazón en el laboratorio, los científicos pueden tener acceso a células humanas adultas sin la necesidad de tomar el tejido directamente de los pacientes. Los científicos pueden estudiar estas células adultas especializadas en detalle para ver qué es lo que no funciona en ciertas enfermedades o para estudiar cómo las células responden a nuevos tipos de posibles fármacos.
Los investigadores están aprendiendo a crear diferentes tipos de células especializadas encontradas en el cuerpo exponiendo las células ESC a diferentes moléculas de señalización y condiciones de crecimiento. Muchos estudios están investigando cómo podrían utilizarse las células ESC para tratar diferentes enfermedades, desde la esclerosis múltiple hasta la ceguera o la diabetes. En el futuro, la esperanza es hacer células ES de otras especies también. Los investigadores también están trabajando en el laboratorio en expandir y perfeccionar los métodos para crear tipos particulares de células adultas a partir de células ES.
Retos y Mecanismos Moleculares de la Diferenciación
Controlar exactamente cómo se diferencian las células madre embrionarias es un desafío muy importante. La diferencia entre un grupo de células madre y un feto en desarrollo radica en una compleja serie de reacciones bioquímicas que transforma las células troncales en tipos determinados de tejido. Existen diferencias entre el funcionamiento de las ESC en ratones y en humanos. Por razones éticas y morales, los investigadores no pueden utilizar las ESC humanas como lo hacen en los experimentos con las ESC de los ratones. Ello hace que tengan que imaginarse cómo funcionan las ESC de humanos de forma más compleja e indirecta. Otro de los retos a los que se enfrentan es el de controlar exactamente cómo se diferencian las ESC en numerosos tipos de células especializadas. Uno de los objetivos principales es encontrar formas de generar grandes cantidades de células especializadas a partir de células madre uniformes y que den resultados fiables, pero esto no será nada fácil.
La complejidad de células y tejidos que forman un cuerpo animal parte de una bola engañosamente simple de células madre embrionarias, el embrión temprano. La forma en que estas células pierden su pluripotencia (es decir, su capacidad para dar lugar a cualquier tipo de célula) y cómo se diferencian en linajes específicos es uno de los misterios de la biología del desarrollo. El embrión vertebrado empieza como una esfera uniforme de células pluripotentes, que pronto se transforma -a través de un proceso denominado gastrulación- en una estructura más compleja con tres capas de tipos celulares diferenciados. Este proceso de diferenciación implica dos etapas clave que pueden superponerse en el tiempo:
- Salida de pluripotencia: Las CME pierden la capacidad para convertirse en cualquier tipo de célula.
- Especificación del linaje: Las CME se diferencian en células del ectodermo, el mesodermo o el endodermo.
Regulación Génica y Epigenética
Durante el desarrollo embrionario se forman varios cientos de tipos de células diferentes del cuerpo humano adulto, a partir de unas pocas células madre idénticas. El potencial de diferenciación de las células es progresivamente restringido en el curso de este proceso, causando cambios en su morfología y funciones. Los mecanismos básicos de control molecular mediante los cuales las células madre deciden en qué tipos de células embrionarias se convertirán, como las del sistema nervioso (neuroectodermo) o las del meso y endodermo (órganos internos o músculos), están siendo descifrados. Esta decisión está regulada por moléculas de señalización embrionaria, como las señales TGFβ y Wnt, las cuales son transmitidas por factores de transcripción que regulan los genes de la familia de factores T-box, como Eomes y Brachyury. Estos factores son los responsables de "activar" los programas de genes de diferenciación para todas las células meso y endodermo y, al mismo tiempo, actúan como represores de genes, previniendo la formación de tejido neural al suprimir los correspondientes programas de genes.
En los organismos, cada tipo celular especializado expresa un conjunto específico de la totalidad de genes presentes en el genoma; son estos procesos de regulación de la expresión génica lo que caracteriza al linaje celular. Si bien la diferencia en los patrones de expresión génica se da en su mayoría por elementos reguladores cis y trans (promotores y enhancers), hay mecanismos para que estos patrones de expresión se mantengan durante muchas generaciones de división celular.
Metilación del ADN y Modificación de Histonas
Uno de los mecanismos más frecuentes para la regulación de la expresión es la metilación del ADN. En este proceso, las enzimas metiltransferasas añaden un grupo metilo sobre los residuos de Citosina ubicados en las islas CpG, evitando el acceso al ADN. Estos tres factores de transcripción (Oct4, Sox2, Nanog), altamente expresados en células embrionarias indiferenciadas, son importantes para el mantenimiento de la pluripotencia. Se ha demostrado que la importancia de estos factores radica en su capacidad de modificar la cromatina, modificando las histonas y metilando el ADN, para permitir o restringir la transcripción de genes. Si bien estos genes son altamente expresados, su capacidad de mantener la pluripotencia requiere un balance cuidadoso, ya que se ha visto que una perturbación puede conducir a diferentes linajes celulares.
Cuando los nucleosomas están firmemente posicionados sobre el ADN evitando cualquier tipo de transcripción, los vamos a denominar heterocromatina. Por el contrario, cuando los enlaces no son tan fuertes y permiten la transcripción, lo denominamos eucromatina. Generalmente la adición de grupos acetilos genera apertura y por ende una mayor transcripción. Este proceso se lleva a cabo sobre residuos de lisina por acetil-transferasas.
Por otro lado, las metilaciones de histonas tienen comportamientos "menos predecibles", ya que según el residuo metilado pueden promover o reprimir la expresión. A pesar de esto, se conocen ciertas metilaciones que reprimen, como la trimetilación de la lisina 27 de las histonas H3 (H3k27me3) o H3K9me (metilación en lisina 3 de la H3). Un factor crítico, Whsc1 (una metilasa de histonas), se ha asociado con la activación de genes maestros de linaje (Gata4, T (Brachyury), Gata6 y Foxa2), lo que lleva a la especificación de células germinales. Cuando se inhibe Whsc1, la salida de pluripotencia se retrasa y las células madre embrionarias de ratón no pueden diferenciarse eficientemente en células del mesendodermo.
Al bloquear todas las PcG (Polycomb group), las células embrionarias no son capaces de diferenciarse en las tres capas germinales. Si bien las proteínas PcG promueven la diferenciación, estas no pueden mantener el fenotipo diferenciado. Mantener estos procesos activos es la función de los genes de la familia Trx (trithorax). Una vez inician los programas de diferenciación celular, las proteínas Trx son reclutadas en las zonas transcripcionalmente activas, donde catalizan la trimetilación de la histona H3 en la lisina 4 (H3K4me3) y también la acetilación de otras histonas.
Una regulación diferencial entre Oct4 y Sox2 conduce a destinos de línea germinal. Del mismo modo, niveles elevados de Oct4 y reducidos de Sox2 promueven destinos mesodermales, ya que Oct4 bloquea destinos neuroectodermales; si aumenta Sox2 y disminuye Oct4 se derivan destinos ectodérmicos. Teniendo todas las células del cuerpo el mismo genoma, los patrones de unión de los Factores de Transcripción y su efecto sobre los genes marcan la diferencia.
Mecanismos Epigenéticos
Vías de Señalización y Morfogénesis
Todos los mecanismos mencionados deben ser activados en conjunto para poder coordinar la formación de un tejido u órgano (morfogénesis). El mecanismo de transducción de la señal es conservado, aunque sus intermediarios cambian según la señal. Un ligando producido por una célula se une a un receptor extracelular de otra célula. Este receptor cambia su dominio citoplasmático y adquiere una propiedad enzimática, esto cataliza una serie de reacciones que amplifican la señal.
Algunos morfógenos relevantes son las proteínas morfogenéticas óseas (BMP), los factores de crecimiento transformantes (TGF) o los factores de crecimiento fibroblásticos (FGF). Otras vías de señalización, como la JAK-STAT, han demostrado ser necesarias para el mantenimiento de pluripotencia en células embrionarias de ratón. El ácido retinoico también ha demostrado que induce la diferenciación en células humanas y de ratón. La señal de Notch cumple papeles importantes en la auto-renovación y el mantenimiento del nicho de células madre. Se ha descubierto que la vía de señalización de Notch está implicada en la formación del trofectodermo junto con Tead4, regulándose conjuntamente sobre el elemento regulador de Cdx2, lo que garantiza el correcto desarrollo del embrión.
La lectura de las cualidades físicas por parte de la célula se basa en el principio de tensegridad, ya que según la fuerza que actúe directamente sobre la membrana celular, esta se deformará e interactuará con componentes del citoesqueleto como la actina, que puede generar una cascada de señalización. Estas señales generan respuesta por proteínas inducidas por tensión, que pueden modificar la cromatina según la tensión mecánica.
Factores Adicionales en la Diferenciación
La enzima QSulf1 actúa en el reloj biológico del embrión para ayudar a las células a responder a una y no otra de la multitud de señales químicas que reciben. La QSulf1 opera sobre las células progenitoras, una forma de células madre ligeramente más avanzadas que tienen ya menos posibilidades de diferenciación. La QSulf1 representa una nueva clase de enzimas cuya función principal es modificar un importante factor de señalización, denominado proteoglicanasa de sulfato de heparina (HSPG). Así se liberan las moléculas señalizadoras Wnt -que están vinculadas a las HSPG en la superficie de las células- que activan los genes reguladores que dan instrucciones sobre diferenciación. La QSulf1 permite a las células embrionarias expresar un gen maestro regulador del músculo denominado MyoD, que a su vez instruye a estas células para que se conviertan en progenitoras del músculo en lugar de células progenitoras de otros tejidos.
Otro descubrimiento importante es la proteína Mbd3, identificada por investigadores de la Universidad de Edimburgo, que es fundamental en el proceso de diferenciación. En ausencia de la Mbd3, las células madre de ratón permanecen en un estado similar al de las células madre embrionarias, lo que sugiere su rol clave para que la célula abandone el estado de auto-renovación y siga la ruta de especialización. Este hallazgo también ha arrojado luz sobre las diferencias entre las células madre humanas y las del ratón, mostrando que las células de ratón sin Mbd3 son capaces de duplicarse sin la proteína LIF, al igual que las células humanas.
Para la clonación de la oveja Dolly, se estudiaron los ovocitos de mamífero y se encontraron presentes ciertos factores de transcripción (como Nanog, Tdgf1, Utf1, Lin28, entre otros) capaces de reprogramar el núcleo, no solo manteniendo su estado de indiferenciación, sino induciendo en núcleos de células diferenciadas una vuelta hacia el estado indiferenciado. El hecho de que estos factores pueden no solo mantener la célula indiferenciada, sino reprogramar su núcleo una vez diferenciada, ha permitido mediante técnicas de laboratorio inducir estadios de pluripotencia (células iPS).
Diferenciación en Otros Organismos
Históricamente, la embriología se ha concentrado en estudiar el desarrollo animal. Por ello, el uso de organismos modelos como Drosophila melanogaster o C. elegans ha cumplido un papel importante en la descripción de estos procesos.
La diferenciación en plantas superiores se produce a partir de las células meristemáticas que son reclutadas para dar lugar a las células maduras que forman parte de los órganos de la planta. Está demostrado que los genes de la familia WOX están relacionados con la organización de grupos de células durante el desarrollo de la planta. Según estudios realizados en el desarrollo de Arabidopsis thaliana y Solanum lycopersicum, en los que se observó la transcripción y función de los genes WOX4, se constató que estos genes están involucrados en el desarrollo de los haces vasculares de la raíz y en el brote de los órganos laterales en ambas especies. Una reducción de la expresión de WOX4 mediante ARN de interferencia en Arabidopsis tuvo como consecuencia plantas de pequeño tamaño, cuyo floema y xilema no se había diferenciado o lo habían hecho dando lugar a conductos más pequeños de lo normal.
tags: #celula #embrionaria #capaz #de #diferenciarse