Las espermatófitas, también conocidas como fanerógamas (Spermatophyta), constituyen un grupo monofilético dentro del reino de las plantas (Plantae). Este linaje abarca todas las plantas vasculares que tienen la capacidad de producir semillas. El término científico "Spermatophyta" deriva del griego "sperma" (semilla) y "fiton" (planta), significando literalmente "plantas con semilla".
La composición de este grupo coincide exactamente con la del antiguo taxón Phanerogamae, lo que lo convierte en un sinónimo de esta división. Las espermatófitas son también denominadas embriofitas sifonógamas, debido a que el grano de polen produce un tubo (haustorial o polínico) para alcanzar el óvulo y facilitar la fecundación. El nombre proviene del griego "embrios" (embrión), "fiton" (planta), "xifos" (tubo) y "gamos" (unión sexual), interpretándose como "plantas con embrión cuya unión sexual ocurre con tubo".
La monofilia de las espermatófitas ha sido ampliamente consensuada por la comunidad científica. Entre las evidencias morfológicas que respaldan esta monofilia se encuentran la presencia de la semilla y la producción de madera, al menos en sus formas ancestrales. La madera es el xilema secundario, generado por el meristemo secundario conocido como cámbium.
Las espermatófitas se originaron a finales del período Devónico, evolucionando a partir de las lignofitas, que ya poseían producción de madera y ramificación axilar, tal como lo evidencia el registro fósil. En la actualidad, las espermatófitas representan el linaje más extenso de plantas vasculares, contando con aproximadamente 270.000 especies vivientes. Un subgrupo significativo de esta diversidad son las angiospermas o plantas con flores periantadas. Otros subgrupos, comúnmente agrupados como gimnospermas, incluyen las cícadas, los ginkgos, las coníferas y los gnetales, todos compartiendo un ancestro común.
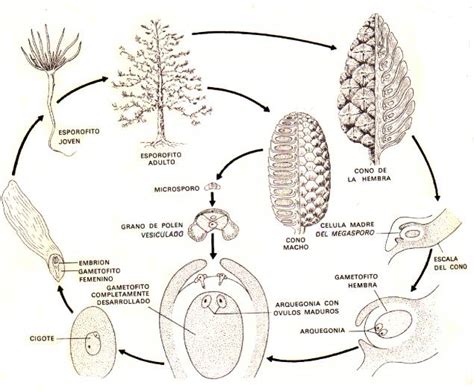
Ciclo de Vida y Estructuras Reproductivas
Al igual que en todas las traqueófitas, las espermatófitas presentan una alternancia de generaciones entre el gametófito y el esporófito. El gametófito es de tipo taloide, mientras que el esporófito está organizado en tejidos y órganos, manifestándose como un cormo con sistema vascular, raíz y vástago. En las espermatófitas, la alternancia de generaciones se da de forma enmascarada, ya que el gametófito se desarrolla completamente dentro de las estructuras del esporófito.
Poseen un ciclo de vida heterospórico, con un gametófito de desarrollo endospórico. Los esporangios se originan en las hojas, denominadas esporofilos cuando son fértiles. En las espermatófitas, las hojas fértiles se encuentran sobre una rama de crecimiento limitado, conocida como braquiblasto. Esto da lugar a una nueva estructura característica: la flor, una rama de crecimiento definido que porta hojas fértiles.
Morfológicamente, una flor puede presentar un eje que se origina en el vástago, con una o dos brácteas, culminando en un receptáculo que alberga los megasporofilos (portadores de megasporas) y los microsporofilos o estambres (portadores de microsporas). Los megasporofilos y microsporofilos, en conjunto, forman los antofilos, lo que explica por qué este taxón también se denomina Antophyta, aunque este nombre no es universalmente recomendado ya que algunos autores lo restringen a angiospermas y gnetales.
Desarrollo de Megasporas y Gametofitos Femeninos
Dentro de la flor, las hojas fértiles que portan megaesporangios (megasporofilos) contienen el megaesporangio. Aquí, tras la meiosis, tres de los productos degeneran, y el esporófito sobreviviente se desarrolla en la única megaspora funcional. A diferencia de otras plantas vasculares, esta megaspora nunca abandona el megaesporangio, una característica distintiva de las espermatófitas.
La megaspora produce internamente el gametófito femenino, el cual puede variar en número de células (entre 6 y muchas) en su estado adulto y tampoco abandona nunca a la megaspora. Este fenómeno se conoce como gametófito endospórico, común en plantas heterospóricas. El gametófito femenino, a su vez, genera uno o varios gametos femeninos, que, como en todas las embriofitas, permanecen dentro de él.
La cubierta del megaesporangio, mediante mitosis, forma los tegumentos protectores del futuro embrión (uno en gimnospermas y dos en angiospermas). Dado que la megaspora no abandona el megaesporangio, este último posee una abertura especializada, la micrópila, para recibir el grano de polen y permitir la fecundación. En las gimnospermas, el óvulo y su micrópila están expuestos; en las angiospermas, están cubiertos por el carpelo.
Formación de Microsporas y Granos de Polen
En la flor, las hojas fértiles que portan microesporangios (microsporofilos) se denominan estambres. En los estambres de las espermatófitas, el saco polínico origina las microsporas. Cada microspora contendrá un único gametófito masculino. El gametófito masculino maduro, envuelto por la pared de la microspora, constituye el grano de polen.
El proceso de polinización implica el transporte del polen hasta las estructuras femeninas preparadas para recibirlo, mediado por agentes externos. En las gimnospermas, el polen se dirige directamente a la micropila; en las angiospermas, se deposita en el estigma del carpelo.
Origen y Evolución de las Espermatófitas
Las espermatófitas descienden de un ancestro caracterizado por la homosporía (un solo tipo de espora y gametófitos bisexuales), similar a los helechos actuales. La heterosporía se originó múltiples veces de forma independiente en linajes no emparentados de plantas vasculares.
En la línea evolutiva que condujo a las plantas con semilla, el esporófito adulto produce meiosis de una sola célula por megasporangio. Mediante el aborto de tres de los productos de la meiosis, el número de megasporas funcionales se reduce a una. Esta única megaspora funcional, en las espermatófitas, se retiene dentro del megasporangio, desarrollando su gametófito femenino y gameta femenina completamente dentro de la generación esporofítica anterior.
Nuestro conocimiento sobre el origen de la semilla se basa en fósiles del Devónico tardío y Carbonífero temprano, denominados "progimnospermas" o "helechos con semilla". La diferenciación entre un tallo principal y ramas laterales ya existía en el linaje de las eufilofitas. La aparición de troncos de gran tamaño con madera similar a la de las coníferas modernas, junto con extensos sistemas de ramas y hojas pequeñas, se observó en el Devónico tardío. La reconstrucción de fósiles como Archaeopteris y otras "progimnospermas" ha sido crucial para establecer el origen de la heterosporía y la producción de madera, concluyendo que ambos precedieron a la evolución de la semilla.
Por ello, es incorrecto clasificar a estos fósiles como espermatófitas, aunque sean ancestros de las plantas con semilla actuales. Análisis de las primeras semillas revelan que se encontraban en "cúpulas" y cada semilla estaba recubierta por una excrecencia de la pared del esporangio, formando una "cámara polínica" especializada para recibir polen, posiblemente asistida por una gota de polinización pegajosa.

Se ha demostrado que el grupo de las gimnospermas vivientes es monofilético y hermano de las angiospermas. Las gimnospermas presentan fecundación simple, a diferencia de la doble fecundación de las angiospermas.
Clasificación Histórica y Filogenia
Durante gran parte del siglo XX, los linajes de plantas con semilla se dividían comúnmente en dos grandes grupos: las cycadofitas y las coniferofitas. Las cycadofitas, incluyendo las cícadas modernas, se caracterizaban por una producción de madera limitada (leño manoxílico) y hojas grandes tipo fronde. Las coniferofitas, como el Ginkgo y las coníferas, poseían madera bien desarrollada y densa (leño picnoxílico), y hojas simples, a menudo en forma de aguja.
Esta distinción llevó a algunos investigadores a proponer que las plantas con semilla se originaron dos veces. Sin embargo, análisis filogenéticos que incluyen linajes vivientes y fósiles generalmente apoyan un árbol filogenético donde la semilla apareció una sola vez. En este árbol, las primeras plantas con semilla se asemejaban más a las cícadas, con hojas grandes y semillas de simetría radial. Fósiles del Devónico-Carbonífero, como Lygniopteris y medulosas, se sitúan en la base de la filogenia de las plantas con semilla, mientras que las coniferofitas forman un clado "platyspérmico" (semillas con simetría dorsiventral).
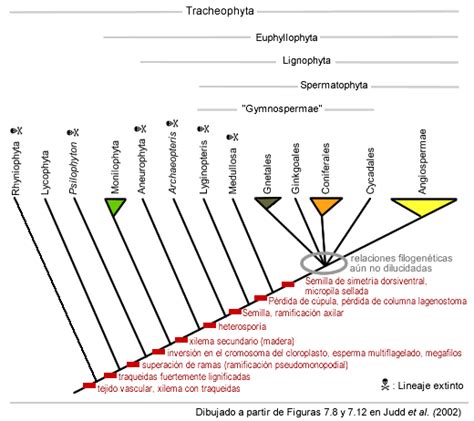
A pesar de los esfuerzos por dilucidar las relaciones filogenéticas de los cinco grupos vivientes de espermatófitas (cícadas, ginkgos, coníferas, gnetofitas y angiospermas), aún existen debates científicos. Se ha descartado que las angiospermas deriven de una gnetofita ancestral, basándose en diferencias morfológicas en los vasos xilemáticos y la estructura floral, así como en hallazgos fósiles que confirman la pertenencia de las gnetofitas al grupo de las gimnospermas.
Grupos Principales de Espermatófitas
Actualmente, existen dos linajes vivientes de espermatófitas: las gimnospermas y las angiospermas. Las gimnospermas fueron abundantes y diversas durante el Mesozoico, quedando hoy alrededor de 130 especies.
Gimnospermas
- Cícadas: Generalmente presentan un tronco bajo y ancho, con xilema secundario limitado y hojas compuestas grandes. Son dioicas, con esporófitos que portan óvulos o estróbilos masculinos productores de polen. Sus semillas suelen ser grandes, a menudo con un tegumento externo carnoso. Características como el polen con tubo haustorial y el esperma gigante multiflagelado pueden ser ancestrales, aunque también poseen caracteres derivados como la pérdida de ramificación axilar y la producción de raíces coralloides.
- Ginkgos: Solo existe una especie sobreviviente, Ginkgo biloba. Esta especie es raramente encontrada en estado silvestre, pero ha sido cultivada y preservada durante siglos.
- Coníferas: Grupo diverso de árboles y arbustos caracterizados por su madera densa y hojas en forma de aguja o escama.
- Gnetales: Grupo de plantas con características intermedias entre gimnospermas y angiospermas, incluyendo plantas como Ephedra y Welwitschia.
Angiospermas (Plantas con Flores)
Las angiospermas son el grupo más diverso y reciente de espermatófitas, caracterizadas por la presencia de flores y la producción de frutos. La clasificación de las angiospermas se basa fundamentalmente en el número de cotiledones presentes en la semilla.
Cotiledones: La Primera Hoja del Embrión
Dentro de la botánica, el cotiledón se define como la hoja primordial de las plantas fanerógamas que se desarrolla durante la germinación de la semilla, constituyendo la primera hoja del embrión. El número de cotiledones es un criterio clave para la clasificación de las plantas fanerógamas.
Características de los cotiledones:
- Son la primera hoja que emerge del embrión.
- Pueden diferenciarse de otras hojas por su tamaño.
- Su número es un método de clasificación vegetal.
- Son capaces de digerir el albumen (tejido nutritivo de la semilla) y proveen nutrientes esenciales para la germinación.
🌱 TIPOS de germinación [Epigea / Hipogea] Según cotiledones
Monocotiledóneas
Las monocotiledóneas son angiospermas que poseen un solo cotiledón en la semilla. Tras la germinación, presentan una única hoja primitiva. Estas plantas no experimentan un verdadero crecimiento secundario, carecen de tronco verdadero y de cámbium, lo que les impide generar madera. Ejemplos típicos incluyen gramíneas (trigo, maíz), lirios, palmeras, tulipanes y orquídeas.
Dicotiledóneas
Las dicotiledóneas son el grupo más numeroso, con embriones que poseen dos cotiledones. Al germinar, generan dos hojas primitivas que nutren a la plántula. Sus hojas pueden presentar diversas formas y bordes. Las ramas de estas plantas están compuestas por anillos anuales de floema y xilema, y son capaces de formar madera. Se conocen alrededor de 170.000 especies, incluyendo rosáceas, leguminosas y rutáceas.
La palabra "cotiledón" proviene del latín "cotyledon", que originalmente designaba a una planta crasa. Linneo, en 1751, le otorgó el significado científico de "hojilla primordial de un embrión".